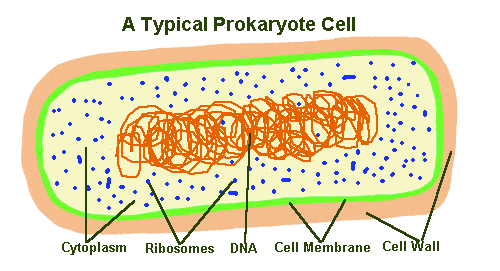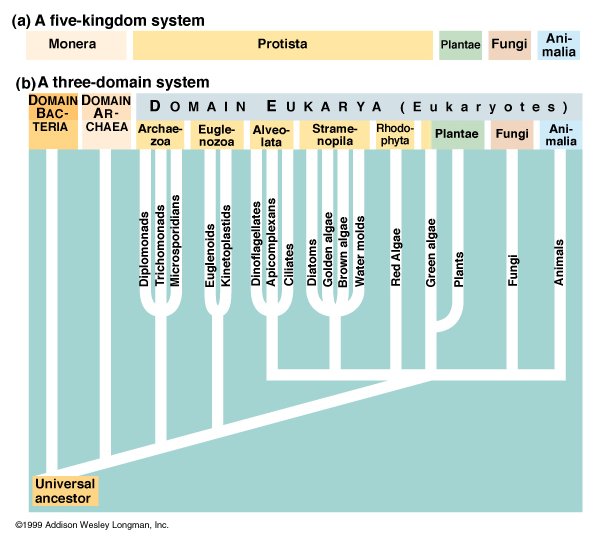
Because the question of which organisms have mental states is an open one, I shall examine case studies that are taken not only from animals, but from various other branches of the "tree of life".
At the present time, there are diverse opinions among scientists regarding what the major groups of living things are, and how they are related. Even the appropriate metaphor for explaining the relationships between living things is a matter for debate - hand? tree? bush? vine? net? ring? What follows is a brief explanation, culled from a variety of sources, mainly collaborative Web projects produced by biologists around the world - BioMedia Associates (2003), The Tree of Life (Doolittle, 2002), the Expert Center for Taxonomic Identification (Hovenkamp, 2002), and the University of California Museum of Paleontology (Waggoner and Speer, 1994, 1998; Kazlev, 2002) - as well as articles by biologists (Doolittle, 2000; Forterre and Philippe, 1999; Margulis, 1989 - 1991) and science reports (Chui, 1999).
Table 2.1 - An overview of some different schemes for classifying living things.
Adapted from a table by M. Alan Kazlev, University of California Museum of Paleontology.
| Two kingdoms (Linnaeus, 1735) |
Five kingdoms (Whittaker, 1969) |
Two domains (used in Margulis' scheme) | Three domains and who knows how many "kingdoms" | |
| Animalia | Animalia | Eukaryotes | Eukarya | Animalia |
| Plantae | Fungi | Fungi | ||
| Plantae | Plantae | |||
| Either classified as Protozoa (Animals) or Algae (Plants) | Protoctista | Alveolata | ||
| Stramenopiles | ||||
| etc... | ||||
| Sporozoa | ||||
| Mycetozoa | ||||
| Euglenozoa | ||||
| etc... | ||||
| Archezoa | ||||
| Formerly classified as plants (bacteria and blue-green algae) | Monera | Prokaryotes | Eubacteria (or bacteria) | (Kingdoms not specified) |
| Archaea | Euryarchaeota
Korarchaeota Crenarchaeota |
|||
|
Definitions of domains:
The term prokaryotes was (and still is) used to describe organisms without a cell nucleus, whose DNA floats freely within the cell (bacteria, etc.) and eukaryotes as organisms whose DNA is stored in a nucleus (animals, plants, fungi, and protoctista). Prokaryotes are now divided into two domains, Bacteria and Archaea, as different from each other as either is from the third domain (Eukaryotes). Archaea are a recently discovered group of microorganisms that can live in extreme habitats such as thermal vents or hyper-saline water. |
||||
|
Definitions of kingdoms used in Whittaker's five-kingdom system (still used by some authors):
1. Monera: includes bacteria, whose cells lack nuclei. Most but not all are single-celled (unicellular) organisms. 2. Protoctista ("first builders"): includes single-celled microbes with nuclei, that were previously labelled protista (protozoa and protophyta), as well as certain multi-celled organisms, such as kelp, that do not belong to the plant, animal or fungi kingdoms. Amoebae, algae, seaweeds, slime moulds, ciliates, diatoms, paramecia and forams belong in this kingdom. 3. Fungi: placed in a kingdom apart from plants, because their life-cycle, feeding habits (they do not photosynthesise) and ecological adaptation are so different. 4. Plants: multicellular organisms which develop from embryos and whose cells contain chlorophyll, enabling them to make their own food. 5. Animals: multicellular organisms that are heterotrophic (feeding on others), diploid (with two sets of chromosomes), and that develop from the fusion/fertilization of an egg and a sperm cell. They follow certain stages during their embryonic development: a morula appears first, then a blastula and finally a gastrula - a hollow sac that forms the embryonic precursor to the digestive tract, by means of which animals ingest nutrients and excrete waste. |
||||
| Note on the Linnean system:
Members of each kingdom were classified in a nested hierarchy (expanded after Linnaeus' death). For example, human beings belong to the phylum chordates (very roughly, animals with backbones), class mammals, order primates, family hominids (humans and great apes), genus Homo and species sapiens. Scientific acceptance of Darwin's theory of evolution allowed biologists to view this scheme as a tree, with descendant branches diverging from a common ancestral stem. However, evidence began to accumulate in the 19th century, following the work of Darwin and Haeckel, that the two-kingdom scheme was too narrow to express the rich diversity of life. The two-kingdom classification gradually fell into disfavour. |
||||
|
Note on the Five-Kingdom Classification:
In 1969, Robert Whittaker proposed his classification of life into five kingdoms: monera (bacteria), protista (now known as protoctista - water-dwelling microbes, such as amoebae), plants, fungi and animals. This scheme is still used by some authors. |
||||
| Note on the Two Domain-Five Kingdom system:
In her book "Five Kingdoms: An Illustrated Guide to the Phyla of Life on Earth" (1987), Lynn Margulis (with Karlene Schwartz) proposed a unified two domain-five kingdom system. In contrast to the usual tree of life metaphor, Margulis used the illustration of a hand, with prokaryotic monera corresponding to the thumb, and the four other eukaryotic kingdoms corresponding to the fingers of a hand. As Stephen Jay Gould notes in his foreword to the book, in this new taxonomy, "the greatest division is not even between plants and animals, but within the once-ignored microorganisms - the prokaryotic Monera and the eukaryotic Protoctista" (italics mine). |
||||
|
Note on the Three Domain system:
Research in the 1990s showed that prokaryotes are far more diverse than anyone had suspected, and a new three-domain classification has emerged. Prokaryotes are now divided into two domains, Bacteria (or Eubacteria) and Archaea, and Eukaryotes are placed in a third domain. |
||||
Because these three domains, the major lineages of the tree of life, are thought to have swapped a lot of their genes in the past (a phenomenon known as lateral gene transfer), mutated several times at the same sites in their genes (saturation), undergone gene duplication (non-orthologous replacement), acquired foreign genes (through eating), discarded different genes (differential gene loss) and evolved at different rates (giving rise to the long branch attraction phenomenon), the rooting of the tree of life remains contentious.
Table 2.2 - Some contemporary views of the rooting of the tree of life
| (1) Prokaryotes, being simpler in their cell organisation, evolved first, and split into bacteria and archaea, while eukaryotes later branched off from the archaea (the standard view until recently). |
| (2) All three domains diverged at the same time from a common source. |
| (3) Eukaryotes represent the root of the tree of life, while prokaryotes are simplified life forms that arose through gene loss and other reductive mechanisms; |
| (4) Eukaryotes arose from a merging of bacteria and archaea. |
| (5) A fourth branch of the tree, now lost, contributed genes to the eukaryotes. |
| (6) There is no unique "tree of life", but rather a tangled thicket of trees (or a net, as Doolittle (2000) describes it): owing to the prevalence of gene-swapping (lateral gene transfer) between different domains of organisms in the past, different genes in the same organism have different family trees, so the same organism can be classified in different ways, depending on which of its genes we examine. Proponents of this view also question the idea that all living things diverged from a common ancestral stock, and suggest that there may be many ancestral roots. |


The upshot of this is that at the present time, it would be very unwise to describe any group of living things as "primitive". For the purposes of this thesis, I shall content myself with describing prokaryotes (bacteria and archaea) as being structurally (not computationally) the simplest cellular life-forms.
The main themes that have emerged from my investigation of sensory capacities in living things are the need for definitional clarity, more careful distinctions between different grades of sensitivity and a greater understanding of the relation between sensitivity and motion in organisms.
What is a sensor?
Definitions for terms such as "sensor", "sensitivity", "sensation" and "sentient" abound in the literature. My definition of "sensor" is adapted from a definition in common use on the World Wide Web, but I have added a condition (in italics):
Definition - "sensor", "sensitive"
A sensor is any device that responds in a specific way to a physical stimulus (e.g. chemicals, heat, light, sound, pressure, motion, flow). A sensor (and by extension, any entity possessing sensors) can be described as sensitive to the stimulus to which it specifically responds.
What is important here is that the response is specific: the mere ability to respond to changing circumstances is an insufficient criterion even for sensitivity, as the response may be a random one.

For instance, a virus is able to respond to changes in its environmental conditions by adopting a bet-hedging strategy: when it invades a host bacterium, it may kill its host immediately by multiplying until the host's cell walls burst (lysis) or it may remain dormant and may confer immunity to infection upon its host (lysogeny). I discuss this case at further length in section 1.1 (case study 1) of the Appendix to chapter 2 part B. The important point here is that the strategy a virus adopts on a particular occasion is a random one, which is triggered by thermal background noise. There is no meaningful sense in which the virus can be said to possess what Dennett (1997) would call sensors, as its response to its environment is non-specific and it encodes no information that would help it to achieve its goals.
S.1 An organism's possession of sensors, which enable it to encode information about its environment and thereby achieve its goals, is a necessary condition for our being able to describe the organism's behaviour according to Dennett's (goal-centred) intentional stance.
S.2 An organism must be capable of encoding and storing information about its environment before it can be said to possess mental states (in particular, beliefs and desires). (Corollary of I.1 and S.1.)
In principle, anything is capable of acting as a sensor: camera film is photosensitive, as are metals which release electrons when exposed to light (the photoelectric effect). The bimetallic strip in a thermostat is a temperature sensor.
Any sensor can be described using Dennett's intentional stance: it is a "micro-agent, ... an utterly minimal intentional system whose life project is to ask a single question, over and over - 'Is my message coming in NOW?' ... - and spring into limited but appropriate action whenever the answer is YES" (1997, p. 108). It is intentionality at this level, Dennett argues, that makes perception possible, and allows an animal to process information about its surroundings.

Any entity possessing sensors can be described as sensitive to the changes its sensors detect. Dennett (1997, p. 84) regards the sensitivity displayed by phototactic bacteria as fundamentally the same as the photosensitivity of light meters in cameras. I would argue that he is profoundly mistaken here: in the former case, but not the latter, the sensors are part of the body of a living individual, which benefits from its ability to sense changes in its environment. It was argued in the previous chapter that there is a fundamental distinction between individual organisms (such as bacteria), which possess both a distinctive formal cause and a final cause (as shown by their possession of a master program that regulates their internal structure, a nested hierarchy of organisation, and dedicated functionality), and the merely extrinsic finality found in current man-made devices, which are assemblages rather than individuals, and which lack a "good of their own".

As it turns out, sensors are found in all cellular organisms, including bacteria, whose sensory capacities I describe in detail, in section 1.1 (case study 2) of the Appendix to chapter 2 part B.
S.3 All cellular organisms (including bacteria) possess sensors that can encode various states of information about their surroundings. Such organisms can therefore be described as sensitive to their surroundings.
Which organisms have true senses?
Should we then say that bacteria possess bona fide senses, or is there a distinction between sensing an object, and being sensitive to it? For that matter, what are senses, anyway?
Some philsophers, such as Aristotle (De Anima 2.11, 424a1 ff.), have argued that there is more to sensing a stimulus than being affected by it. If they are right, an organism's sensitivity to a stimulus need not imply that it can sense the stimulus.
Aristotle maintained that senses are not found in all living things. Aisthesis - which in his writings "is capable of bearing the meanings both of sensation and of perception" (Lawson-Tancred, 1986, p. 78; see also Sorabji, 1993, pp. 8, 15) - is found in animals, and only animals (De Anima 2.2, 413b1ff; 3.12, 434a30; 3.13, 435b1). Aristotle stresses that senses exist for a practical, teleological reason: they are discriminative capacities (De Anima 3.9, 432a16), which enable animals to survive. Without these capacities, animals cannot avoid danger or acquire what they need (De Anima 3.12, 434b - 1ff). This is particularly true of animals that move around: "If any body that travels did not have perception, it would be destroyed and so not achieve nature's function by reaching its purpose" (De Anima 3.12, 434a33-34). Since bacteria also travel, any distinction that may be drawn between the sensory capacities of animals and the sensitivity exhibited by bacteria surely needs to be carefully justified on philosophical grounds. Does Aristotle's definition of sensation offer such a justification?
Aristotle argued that there is more to sensing an object than merely being affected by it:
For perception is being affected in a certain way. Thus the active thing [the sense object - V.T.] makes that [the sense organ - V.T.] which is potentially like it, like it in actuality...[T]he sense is the recipient of the perceived forms without their matter, as the wax takes the sign from the ring without the iron and gold... And it is also clear why it is that plants do not perceive, though they have a psychic part and are in some way affected by the touch-objects. After all, they become cold and hot. The reason, then, is that they do not have a mean, nor such a principle as can receive the forms of the sense-objects, but are affected by the matter as well (De Anima 2.11, 424a1; 2.12, 424a17-19, 424a34-424b3, italics mine).
What Aristotle seems to be arguing is that although a living thing which senses an object (with one of its sense organs) is altered by that object, it is not altered by taking that object into itself, but rather by taking on the object's form, without its matter. Rephrasing Aristotle's insights in contemporary terminology (which is etymologically rooted in the form-matter distinction), we might say that for Aristotle, to be able to sense or perceive an object means, roughly, to be a living thing with a sensory organ that can encode information about that object.
There remains the question of what Aristotle meant by saying that aisthesis is "a kind of mean of the opposition in the sense-objects, and thus a judge of them" (De Anima 2.11, 424a1ff). Later, he argues that to perceive A and B (e.g. white and black) the sense-organ must be neither in actuality but both in potentiality. How does this notion of a mean tie in with the requirement that sensory organs be capable of receiving forms without matter? I would suggest that the key lies in Aristotle's remark that "it is the mean that judges" (De Anima 2.11, 424a6). Since aisthesis is a discriminatory capacity, the sense organ needs to not only be able to encode information about its object, but also be separable from the information that it encodes about its object - i.e. it is not always actualised in the same way, and may even be capable of existing in an inactive state. Our eyes can be said to sense colour, only because they do not always see any given colour: at night, they see nothing at all. (Aristotle's remarks on vision are unfortunate; he regards white and black as "two ends of the scale" (De Anima 2.11, 424a7-8), whereas black is, in reality, the absence of light - precisely the kind of mean he is looking for.) A sensory deprivation tank is one way of illustrating the notion of a mean: here, we have a perfect separation of sense organs from the sensory information they encode, as none of the sense organs is being activated.
Thus for Aristotle, an organ of a creature that senses is "informed" by the stimulus it detects (that is, receives the form of its sense-object without the matter), and is also informed about the stimulus. Specifically, because the sensitive organ can be actualised in any one of a number of ways, depending on the form it receives, it can serve as a "mean" between the various sense-objects it is capable of receiving.
In contemporary jargon, what Aristotle is saying is that an organism which can sense should possess organs (sensors) that can receive, encode and store information relating to the stimulus, which corresponds to different states of the stimulus. Sensors that merely detect the presence or absence of the stimulus are not truly informative.
On my interpretation of Aristotle's account, there are thus two ways in which sensitivity in an organism can fail to qualify as a bona fide sensory capacity: either the actual state of the object sensed may not be formally encoded as information, or the sensor may be unable to represent different actualisations or states of its object (e.g. different temperatures). A chemical reactant fails the first of Aristotle's conditions: it receives the matter as well as the form of its object. A binary sensor fails the second condition: because it is unable to represent different actualisations or states of its object, it cannot properly be said to sense it.
The distinctions drawn by Aristotle are by no means trivial ones. The question is: does the sensitivity exhibited by bacteria measure up to Aristotle's criteria for sensory capacities?
Although Aristotle was not aware of the existence of microbes, we now know that even the humblest bacteria possess chemical sensors that satisfy his criteria for possession of senses, thereby rendering obsolete his restriction of senses to animals. I argue this point at further length in section 1.1 (case study 2) of the Appendix to chapter 2 part B. Plants also possess sophisticated sensors that can convey a wealth of information about environmental stimuli which I discuss in section 1.3 (case study 2) of the Appendix to chapter 2 part B. Senses, as Aristotle understood them, appear to be a universal feature of cellular organisms.
What kind of senses exist in organisms?
However, it would be wrong to conclude that the term "sense" has the same meaning when applied to all these organisms. Organisms' sensors differ vastly, not only in their degree of sophistication, but also in the way in which they move organisms to obtain their goals (Cotterill, 2001). I discuss this at further length in section 1.3 of the Appendix to chapter 2 part B. Bacteria, for instance, can only obtain information about their surroundings by continually probing them, through the propulsion provided by their flagella. Thus in bacteria, locomotion is prior to the reception of sensory information, whereas in eukaryotes (plants, animals, fungi and microbes whose cells have nuclei) it is the other way round: information received by these organisms' sensors makes them move towards their goals. Cotterill (2001) argues that this difference is philosophically significant. Additional distinguishing criteria between organisms which Cotterill considers relevant include:
- the presence of dedicated receptor cells (as opposed to receptor molecules on the surface of the organism);
- the presence of a neuron-based movement mediator which is capable of activating different motor programs in response to environmental feedback;
- the presence of reflexes, which produce motor responses independent of the creature's internal state; and
- the presence of a sensory processor which allows an organism to form correlations between various kinds of sensory inputs.
Definition - "sense"
On a broad definition of "sense", any organism possessing sensors that can encode and store information relating to a stimulus, which corresponds to different states of the stimulus, can be said to sense the stimulus. On a narrower definition, the verb "sense" can be restricted to organisms possessing sensors with the ability to:
- initiate movement in an organism towards a goal;
- activate different motor programs in response to environmental feedback;
- activate fast-action responses (reflexes) to stimuli, which are independent of the organism's internal state;
- in multi-sensory organisms, form correlations between different kinds of sensory input.
S.4 On the broad definition used above, all cellular organisms (including bacteria) can be said to possess senses. In eukaryotes (but not prokaryotes), sensors initiate movement towards goals; and reflexes appear to exist in two species of coelenterates, as well as all "higher" phyla of animals.
It should be clear that the attribution of senses to various kinds of organisms is not a univocal one.
What kind of senses are required for intentional agency?
I discuss the philosophical implications of bacteria's "locomotor-driven" senses at further length in section 1.2 of the Appendix to chapter 2 part B. Although they continually probe their surroundings by making random tumbling movements, the movements they make towards attractive stimuli are quite different, consisting of directed "runs" along chemical gradients. Such directed movement could conceivably qualify as "movement started by the object of desire" (Aristotle, De Anima 3.10, 433a16), as it is subsequent to the act of sensing the attractant. Thus there appears to be no reason in principle why an intentional agent could not possess the kind of locomotor-driven senses that bacteria have. Cotterill (2001) makes an excellent case that consciousness requires more advanced senses, but the kind of agency I am considering here may not be conscious.
We might suppose that a bacterium desires the chemicals it is attracted to, and even entertains simple beliefs about them, of the form "food-here-now".
On the other hand, there seems to be no good scientific reason for employing mentalistic terminology to describe a bacterium's ability to sense objects. The mind-neutral goal-centred intentional stance described above, which explains the organism's behaviour in terms of its information about its goals, appears to be adequate, thereby rendering redundant any attribution of beliefs and desires to it. There is no extra explanatory work for an agent-centred stance to do.
S.5 The possession by an organism of sensors which encode information about its environment is an inadequate warrant for saying that it is capable of cognitive mental states.
As we have seen, the senses found in different kinds of organisms differ vastly in their level of sophistication. Even if the sensory capacities of bacteria do not need to be explained in terms of beliefs, could an agent-centred stance be the most appropriate way of explaining the more sophisticated senses found in "higher" animals, which can initiate goal-directed movement, activate motor programs, trigger reflexes and correlate sensory inputs?
I would argue otherwise. From a teleological perspective, the primary significance of an organism's ability to sense is that the act of sensing makes it aware of the presence of one of its built-in ends (e.g. food), in order that that it can pursue this end. (Senses may also make an organism aware of means to an end, but that is not their essential function.) However, there seems to be no scientific advantage in invoking beliefs, desires and intentional agency to explain the pursuit of an end as such; a goal-centred stance, which construes the behaviour as information-guided pursuit of a built-in goal, accounts for the behaviour perfectly well.
Lastly, I do not wish to contest the obvious fact that in human beings (and some non-human animals), the act of sensing an object normally gives rise to certain beliefs about that object. However, the question at stake here is not whether sensing is normally accompanied by the formation of beliefs, but whether the mere possession of senses by an organism is a guarantee that is capable of forming beliefs. The fact that some animals with senses also happen to have beliefs sheds no light on this question.
S.6 The fact that an organism can sense objects in its environment is an inadequate warrant for saying that it is capable of cognitive mental states, even if the organism's senses are of the sophisticated kind found only in "higher" animals.
In the interests of clarity, we need to distinguish between various kinds of memory, as we did with senses. The following broad definition is adapted from a definition of memory used by Microsoft (http://www.microsoft.com/office/newtosite/glossary.asp):
Definition - "memory"
The term memory refers to any capacity for storing information.
Defined in this way, memory is a very common feature of both natural and man-made systems: indeed, it can be said to occur in "any system that has structures ... that can persist for a long time and affect the behaviour of the system" (Wolfram, 2002, p. 823).

The simplest kind of memory found in organisms is a kind of chemical memory found in bacteria, which use it to search for food. Because bacteria are too small to detect any changes in the concentration of nutrients from one end of their body to the other, their only way of deciding which way to move in their search for food is to rely on a very short-term memory mechanism, in which they move around randomly, sample the concentrations of chemicals in their environment at regular short intervals, and compare the current concentration of attractant chemicals in its environment with the concentration during the last measurement. If there is an increase in the concentration of attractants, the bacteria will keep moving in the same direction.
M.1 All cellular organisms possess some kind of memory capacity, which enables them to detect changes in their environment.
We can describe the behaviour of these bacteria in two ways. We could adopt an agent-centred intentional stance, and say that they are exhibiting "purposeful movement" and are searching for food, on the basis of what they remember. Or, we could adopt a mind-neutral, goal-centred intentional stance (the bacteria, in response to stored information, are moving along a chemical gradient toward their goal).
There are two good reasons for favoring a goal-centred stance. First, the three-second bacterial memory is severely limited, for reasons I describe at further length in section 2.1 of the Appendix to chapter 2 part B:
it is binary (the bacterium simply decides whether to keep tumbling randomly or keep moving in its direction),relative (the bacterium does not remember absolute concentrations of attractants, but changes in concentrations) and
ahistorical (no chemical record is kept of the magnitudes of the various concentrations at different times - instead, the bacterium simply compares its present circumstances with its situation a few seconds earlier).
Second, the crucial test that a mentalistic explanation must satisfy is that it allows us to make new or better scientific predictions.there appears to be no scientific advantage in describing the foraging behaviour of bacteria as mindful and interpreting it in terms of beliefs and desires. A goal-centred intentional stance appears to explain the behaviour of the bacteria equally well.
M.3 The chemical memory of bacteria can be adequately described using a goal-centred intentional stance.
M.2 The existence of memory in an organism is not a sufficient ground for ascribing cognitive mental states to it.
Other kinds of memory in organisms
In addition to the simple chemical memory found in bacteria, psychologists have identified several other kinds of memory in organisms, although controversy continues regarding the classifications used. Roediger, Marsh and Lee (2002) provide a useful summary of the current literature in the field. The distinction proposed by Ryle in 1949 between declarative memory and procedural memory - roughly, between "knowing that" and "knowing how" - is still widely invoked, although some recent authors refer to the latter simply as non-declarative memory. Tulving (1972) has suggested that there are two forms of declarative memory: an episodic memory for particular events (e.g. "Where did you go on vacation last summer?"), which involves accessing the time and place of their occurrence, and a semantic memory for general facts about the world (e.g. zebras have four legs). The distinctions are not as clearcut as one would like: it seems that most if not all memory tasks have some procedural component, although some procedural tasks (e.g. tying one's shoe-laces) require no declarative component (Roediger, Marsh and Lee, 2002, pp. 5-6).
M.4 The distinction between procedural (non-declarative) and declarative memory - "knowing how" versus "knowing that" - appears to be a fairly robust one.
In evolutionary terms, procedural memory is thought to be the oldest and is said to be shared by all animals (Tulving, 1985). However, I present reasons for doubting that sponges and cnidaria (coelenterates) possess this kind of memory in section 2.3 of the Appendix to chapter 2 part B.
Although procedural memory is commonly held to include capacities such as classical conditioning, motor skill learning and complex (skill-based) problem-solving (Roediger, Marsh and Lee, 2002, p. 5), the term "procedural memory" remains poorly defined in the literature. I review a selection of definitions in section 2.2 of the Appendix to chapter 2 part B. The most common item listed in these definitions is "memory for skills", but the term "skill" is vague: does it refer to motor skills, perceptual skills, cognitive skills or all three? Memory for habits is also listed in one definition as a form of procedural memory. While there is general agreement that any organism undergoing classical conditioning (which I discuss in a later section) acquires a new skill, the foregoing definitions leave it open as to whether habituation should count as the acquisition of a habit.
M.5 Procedural memory remains poorly defined in the scientific literature.
I discuss which organisms are capable of procedural learning in section 2.3 of the Appendix to chapter 2 part B. I conclude that there are no credible reports in the literature of habituation in bacteria or archaea, let alone classical conditioning. I also conclude that claims in the literature (see Hennessey et al., 1979, pp. 417-423; Abramson et al., 2002, pp. 175-176) that plants and protoctista are capable of classical conditioning - which is a form of procedural learning - have yet to be properly demonstrated (in the case of plants) or replicated (in the case of protoctista), and that further research is needed.
The assessment of declarative memory in animals is a challenging task, as they cannot verbally declare what they remember. The procedure normally used by scientists to assess animals is the delayed non-matching to sample (DNMS) tasks, in which animals are presented with a sample object and then after a delay interval are asked to choose between the sample object and a novel object. Choosing the novel, non-matching object is the correct choice. The demonstrated ability of primates, rats, pigeons (Young and Wasserman, 2001) and even honeybees (Giurfa et al., 2001) to perform these tasks indicates that they possess some form of semantic memory. Further documentation for other species is supplied in section 2.3 of the Appendix to chapter 2 part B.
Episodic memory is considered as the most recent form of memory, and its occurrence in non-human animals is still disputed, although there is evidence that western scrub jays possess an episodic-like memory (Shettleworth, 2001; Clayton et al., 2003; Emery and Clayton, 2004).
M.6 Procedural memory appears to be common to all animals and possibly some other eukaryotes, but does not occur in prokaryotes. Declarative semantic memory is found in mammals, birds and some insects. The existence of episodic memory in non-human animals remains unproved.
Other distinctions have been made in the literature between explicit memory (intentional or conscious recollection of information) and implicit memory (memories people are not consciously aware of, but that can affect their subsequent performance and behaviour), conscious and unconscious memory, voluntary and involuntary retention, and retrospective and prospective memory. As my concern in this chapter is with the conditions for agency rather than consciousness, I shall not discuss these distinctions here.
The question we have to address is: what kind(s) of memory, if any, does an organism need to possess before we can say that its behaviour is most appropriately described by an agent-centred rather than a goal-centred intentional stance?
Is memory required for intentional agency?
One might argue that no memory is required, as we can perfectly well conceive of an organism with conscious feelings but no memories. However, this objection is irrelevant for two reasons: conceivability does not imply possibility, and our concern here is with agency, rather than consciousness or feelings. The question we should ask instead is whether we could ever be justified in using an agent-centred stance to describe the behaviour of an organism with no memory capacity.
Supposed cases of individuals who have lost all their memories, but still have beliefs and desires, turn out to be nothing of the sort. If we look at the most celebrated example in the psychological literature, the individual known as H.M., it turns out that although he has completely lost his episodic memory of events since his operation, and his semantic memory appears to be frozen in the 1950s (with a few exceptions), his procedural memory remains intact (Rorden, C. University of Nottingham, Course C81HBB (Neuropsychology), lecture 2, http://www.psychology.nottingham.ac.uk/staff/cr1/nsy1b.pdf). Likewise, patients with Korsakoff's syndrome are incapable of forming declarative memories but retain the ability to form procedural memories.
To see why memory in an organism is an essential condition for attributing beliefs and desires to it, I would invite the reader to consider the question of whether there could be an organism whose beliefs were only about its biological ends - in other words, an organism that was capable of entertaining beliefs of the "food-here-now" variety, but unable to form other kinds of beliefs. At first blush, it might seem that the answer is yes: after all, such an organism would certainly be capable of exhibiting behaviour which manifested its desires for its own built-in biological ends, as well as its beliefs about those ends (as stipulated in conclusion I.4).
However, such a description would be scientifically redundant: we would not need to invoke an agent-centred intentional stance to explain the behaviour of such an organism, as a goal-centred stance would do the job just as well. There is no way in principle of behaviourally distinguishing an organism which only has beliefs about its ends from an organism lacking beliefs and possessing only built-in goals, as neither organism (by definition) is capable of forming beliefs about new and better means of achieving its ends.
The case we have been considering is a hypothetical species of goal-seeking organism which never manifests a capacity to remember at any stage of its development. Such an organism lacks what might be called a "natural capacity" to form memories: that is, the internal program that directs its biological development does not encode for the creation of structures with a memory capacity. However, if there were a hypothetical individual possessing a mind, that completely lost its capacity to remember, due to physical deterioration or trauma, but displayed a rich behavioural repertoire consistent with its holding beliefs and having desires, then we might be inclined to say that the impaired individual still possessed some mental states, despite losing its memory.
The foregoing argument suggests that there is a vital teleological link between memory and agency. Memory enhances an individual's ability to realise its own good, by conferring new capacities on it. These capacities enable the organism to realise its built-in ends through different means - new patterns of behaviour. That is what memory is for. Without memory, an organism cannot change its patterns of behaviour.
Beliefs about ends ("food-here-now") are thus unintelligible in isolation; their ascription is redundant unless the individual is capable of learning new ways of obtaining the objects of its desire - in other words, new and useful skills. (The fact that some skills acquired confer no benefit on their possessor in no way weakens the point being made here, that the teleological purpose of memory is the acquisition of useful skills.)
Why is procedural memory required for intentional agency?
What the foregoing argument implies is that imputing beliefs and desires to an organism is warranted only if it possesses some form of procedural memory. This is a strong conclusion: it entails that a hypothetical entity which lacked the ability to acquire new skills but was nevertheless able to learn new facts about the world (semantic memory) or recall events from its past (episodic memory) would still not qualify as being an agent. Why should this be so?
I contend that procedural memory is the most fundamental form of memory relating to agency, and that the intrinsic finality of other forms of memory is derivative upon that of procedural memory. None of the information (whether generic or tied to a specific time and place) that an individual remembers will benefit it unless it is capable of varying its patterns of behaviour in a way that promotes its own well-being. To do this, it must acquire new skills.
Thus the major cognitive limitation of an organism lacking memory capacity is that it would be unable to acquire new patterns of behaviour, as new patterns would have to be stored or encoded somewhere in the organism. How could scientists ever infer that an organism had cognitive mental states, if it was unable to acquire new patterns of behaviour? I shall re-visit this conclusion below, in my discussion of Conclusion F.2, where I shall endeavour to supply it with a firmer basis.
M.8 There can be no scientific or philosophical justification for attributing beliefs and desires to an organism lacking procedural memory. In other words, procedural memory is a necessary condition for the attribution of mental states.
M.9 As there have been no credible claims that prokaryotes (bacteria and archaea) possess any kind of procedural memory, we can assume that they do not have beliefs or desires.
M.7 There can be no scientific or philosophical justification for attributing beliefs and desires to an organism lacking memory. The existence of memory capacity in an organism is a necessary condition for ascribing cognitive mental states to it.
Is procedural memory sufficient for intentional agency?
Even if the possession of procedural memory is a necessary condition for the attribution of beliefs and desires to an organism, that does not make it a sufficient one. From a scientific standpoint, there appears to be no reason why an organism's acquisition of new skills should be better accounted for in terms of its beliefs and desires, rather than the information it possesses in order to achieve its goals.
M.10 Procedural memory is not a sufficient condition for the attribution of mental states.
The upshot of our enquiry into memory is that the existence of a memory for new skills in an organism is a necessary but not a sufficient ground for ascribing cognitive mental states to it.

Does modifiable behaviour warrant the ascription of mental states to organisms?
Indirect, modifiable behaviour is a universal feature of cellular organisms (Kilian and Muller, 2001). When an organism's sensor detects a chemical, it activates a chain of chemical reactions, each of which is reversible. Thus the coupling between stimulus and response is indirect. The coupling is also modifiable: an organism's response to a stimulus may be altered by the presence (or absence) of another stimulus.
F.1 Modifiable behaviour occurs among all cellular organisms. Specifically, in any cellular organism, the reaction to a stimulus is always indirect and modifiable (through the addition or removal of other stimuli).
Some cognitive scientists (cited in Di Primio, Muller and Lengeler, 2000, pp. 4, 7) regard the occurrence of indirect, modifiable stimulus-response coupling as evidence for limited cognition in organisms. I would like to suggest that scientists who regard modifiable behaviour as a sign of cognition may have been misled by an ambiguity in the word "modifiable": it may mean "responsive to external changes", or it may mean "not governed by a fixed rule or pattern". The word "modifiable", understood in the former sense, has no mentalistic connotations. Only in the latter sense does it imply the kind of flexibility that might indicate the presence of cognitive mental states.
In the opposite camp, Kilian and Muller (2001) contend that the occurrence of indirect, modifiable behaviour per se does not warrant the attribution of cognitive states to organisms, as this kind of modifiable behaviour can be fully explained as the resultant (net effect) of fixed, pre-existing patterns of behaviour within the organism itself.
It is of course true that fixed patterns of behaviour may be accompanied by mental states in an organism. For instance, the sight of a predator may trigger a number of fixed behaviour patterns in its prey, and these patterns may be automatically accompanied by the belief that a predator is present and the desire to escape from the predator. But the question at stake here is not whether fixed patterns of behaviour may be accompanied by beliefs and desires, but whether these patterns are sufficient by themselves to warrant the ascription of beliefs and desires. As I take it, the point of Kilian and Muller's argument (which I discuss at further length in section 3.1 of the Appendix to chapter 2 part B) is that there is nothing about fixed patterns of behaviour per se - even when the coupling between stimulus and response is indirect and modifiable - which requires us to explain their occurrence in an organism by ascribing mental states to it.
F.2 Behaviour by an organism which conforms to a fixed pattern or rule is not a sufficient warrant per se for ascribing cognitive mental states to that organism, even if stimulus-response coupling is indirect and modifiable (by the addition or removal of other stimuli).
Although the term "fixed patterns of behaviour" is widely used, it is seldom defined. Kilian and Muller (2001, p. 3) are therefore to be commended for highlighting two features that can help us to define the term: functional specificity and goal specificity. (Note: a similar term, "fixed action pattern", is used in a technical sense by ethologists to describe behaviour which continues in the absence of a stimulus; to avoid confusion, I shall refrain from using the term "fixed action pattern" in this chapter.)
I would like to propose the following simple definition:
Definition - "fixed pattern"
We can mathematically represent a pattern of behaviour in an organism by an output variable (say, z). A fixed pattern of behaviour can be defined as a pattern where the value of the output variable z remains the same, given the same values of the input variables.
A fixed pattern of behaviour presupposes that the function (or more generally, combination of functions) governing the behaviour of the output variable, the number and identity of its input variables and the program instructions governing the function's conditions of activation (or invocation) remain the same. All of these variations can be considered as changes in the suite of programs governing the behaviour of an organism. (These programs should not be confused with the biological "master program", discussed in chapter one, which determines an organism's structure and the interactions between its components.)
Although some authors (Godfrey-Smith, 2001; Carruthers, 2004) regard behaviour that varies over the course of time as evidence of flexibility, I maintain that nothing in the above definition precludes us from using time-dependent historical variables, or even derivatives such as rates of change, as inputs to the value of the function determining the output variable. On my account, behaviour that varies over time may still be quite inflexible. In section 3.2 of the Appendix to chapter 2 part B, I explain why specific instances of flexible behaviour cited by the above authors are better viewed as cases of fixed behaviour.
Another implication of my definition is that even in a fixed pattern, the value of the output variable z may be determined by a combination of two or more different functions, depending on the values of the inputs. (I provide a simple mathematical example in section 3.3 of the Appendix to chapter two part B.) However, because the program has not changed, we can still describe the overall pattern as fixed.
The mathematical reason why a response to a stimulus that is "modifiable" by the addition (or removal) of another stimulus may still be the result of an underlying fixed pattern of behaviour is that adding or removing a stimulus can simply be represented as changing the value of an input variable for one of the functions that determines the value of the output (or response).
The role of fixed and flexible behaviour in the life of organisms
It should be clear from the above definition that fixed patterns of behaviour constitute only a subset of all possible instances of programmed behaviour. An organism's behaviour may be programmed, but the program may change over time - whether because of new patterns of output (i.e. new functions), new kinds of input (new parameters) or new conditions under which the output patterns are generated (new program instructions). Such changes constitute what I would define as "flexible behaviour".
Definition - "flexible behaviour"
If the program governing an organism's behaviour changes over time, such that the value of an output variable z is no longer the same for the same inputs, whether because of a change in the function(s) which define the value of z, or the parameters of the function(s), or the conditions in the program under which the function(s) are invoked, then the behaviour described by z is flexible.
F.3 The occurrence of flexible behaviour in an organism is a necessary condition for the warranted ascription of cognitive mental states to it. (Corollary of Conclusion F.2.)
The distinction drawn between fixed and flexible patterns of behaviour might suggest that we can divide the world into mindless individuals whose patterns of behaviour are utterly rigid, and individuals that can behave flexibly and are therefore eligible candidates for having minds. However, such a dichotomy is both simplistic and profoundly mistaken, as it overlooks a more fundamental division between entities whose patterns of behaviour are internally regulated (by a master program of the kind described in chapter 1) and intrinsically ordered to the benefit of their possessor, and those entities whose patterns lack internal regulation and intrinsic finality. Only entities of the former kind can be said to be alive, with a good of their own. Without the property of life, we could not even consider them as candidates for mental states (see Conclusion B.2).
Cognition, at least in Nature, can exist only in organisms that are able to live without it... All basic bodily functions are controlled automatically at the level of physiological reglation. Essential action patterns are innate... (Strube, 1998, pp. 2, 12).
In other words, flexible behaviour is built upon a supporting bedrock of fixed patterns of behaviour, which organisms require in order to survive.
Why memory matters for having a mind
Conclusion F.2 above allows us to articulate more clearly the basis for our proposal (Conclusion M.7) that the existence of memory capacity in an organism was a necessary ground for ascribing cognitive mental states to it. An organism lacking a memory capacity could only exhibit fixed patterns of behaviour, which do not warrant the ascription of cognitive mental states. A goal-centred intentional stance is sufficient to explain the organism's behaviour.
Is flexible behaviour common in nature?
Where is flexible behaviour found in nature? Over the course of time, evolution may modify information pathways in unicellular organisms. Mutations which alter the DNA or RNA of these organisms may give certain individuals a selective advantage. The evolution of a lineage of organisms may bring about changes in the genetic programs regulating their behaviour, but as these changes do not take place in a single individual, they cannot be treated as evidence of mental states. Kilian and Muller (2001) characterise this as evolutionary adaptation or "phylogenetic 'learning'" (2001, p. 3), which they distinguish from true learning, on the grounds that the changes in behaviour are not "individually acquired during the life-time of the cell" (2001, p. 2). It has been argued above (conclusion B.4) that mental states cannot be meaningfully ascribed to lineages of organisms, but only to individual organisms.
However, even "simple" organisms such as bacteria can also be very adaptable on an individual level. In section 3.4 of the Appendix to chapter 2 part B, I focus on three forms of behaviour - cellular regulation, phenotypic plasticity and gene-swapping - which are commonly cited in the literature, and which might be regarded as the "most promising" candidates for truly flexible behaviour in bacteria. I conclude that only the last of these is truly flexible, and that because it is externally rather than internally generated, it should not be taken as evidence for mental states.
Bacteria possess numerous forms of cellular regulation, the most versatile of which I consider to be the behaviour of bistable switches, where the switch appears to learn a new pattern as its setting fluctuates from "going up" to "coming down". After carefully examining this behaviour, I conclude that it is not truly flexible, and that it is best explained in terms of the memory possessed by bacteria.
Perhaps the best-known instance of phenotypic plasticity in bacteria is the way in which the lac operon is regulated in E. coli. An operon may exist in one of two regulatory states: ON or OFF. Under certain circumstances, the lac operon allows E. coli to use lactose as an energy source, and break it up into its constituent sugars: galactose and glucose. I describe the factors controlling this behaviour and conclude that it does not qualify as truly flexible, as the mechanism (i.e. the relevant program statements) governing the expression of the operon does not vary over time; only the environmental conditions do (i.e. the concentrations of different chemicals). These changing conditions correspond to changes in the values of the input variables.
That leaves us with gene-swapping. One interesting and well-studied example of gene-swapping, which I discuss in detail in section 3.4.3 of the Appendix to chapter 2 part B is the ability of bacterial cells to acquire genes from other bacteria which confer resistance to mercury, a naturally occurring environmental toxin. As the diagram shows, bacteria also employ gene-swapping to transfer resistance to antibiotics. Gene-swapping is a common occurrence among bacteria - indeed, it appears to be a universal trait of organisms, as illustrated by the frequency of lateral gene transfer between different branches of the tree of life. I claim that gene swapping meets all the requirements for truly flexible behaviour. The acquisition of new genes by an organism corresponds to a change in the program statement governing its response to its environment, as well as the acquisition of new functions. Since gene-swapping occurs in all organisms, we can regard the following conclusion as established:
F.4 All organisms exhibit flexible behaviour, to some degree.
Does flexible behaviour warrant the ascription of mental states to organisms?
Since flexible behaviour is often regarded as a hallmark of cognition, it would be tempting to conclude that cognition occurs in all organisms. However, I contend that it is simplistic to equate flexible behaviour with cognition, as flexible behaviour may be externally rather than internally generated. Even if organisms can acquire new information transfer paths (i.e. new cellular program instructions) that enable them to process stimuli differently, they may still lack an in-built mechanism for acquiring information that allows them to modify their response to a stimulus.
I suggest that gene swapping should be regarded as a random exchange of information-processing mechanisms between organisms, whereby they acquire new cellular program instructions. In gene swapping, the process of program modification is externally driven. An organism is incapable of modifying its own program: it can only acquire a new set of pre-packaged instructions from an outside source: another organism. Without this external source, the organism is incapable of changing its response to a stimulus, which remains fixed. In other words, the flexibility conferred by gene swapping is of a very limited sort.
Once the bacterium has acquired the instructions that alter its response to a new stimulus, its information processing pathway remains the same until another "gene-swap" occurs. In the meantime, functional behaviour is still rigidly linked to its goal, as the link between sensors and effectors is still governed by the molecules produced by the cell. One could say that after the swap, the bacterium has simply acquired a new, more adaptive kind of behavioural rigidity.
Internally driven flexible behaviour, on the other hand, arises from a mechanism within organisms, which enables them to "tailor their own responsive dispositions to their particular surroundings" (Beisecker, 1999, p. 298, italics mine). In other words, an internal mechanism for generating flexible behaviour allows an organism to change its pattern of responding to a stimulus without receiving program instructions from outside. Since the change is internally driven, we can say that the organism learns to do something new. When an organism learns to change its response to a stimulus, the change in its behaviour is caused by the stimulus, without the organism having to acquire any new program instructions from the stimulus.
Another feature of gene-swapping which precludes it from being a cognitive (or belief-driven) adjustment to a stimulus is the fact that the change in response bears no relation to the stimulus itself. Indeed, the stimulus (mercury) is incapable of modifying the organism's response to it. Whenever gene-swapping alters the response of an organism (e.g. a bacterium) to a stimulus (e.g. mercury), the changes are caused by foreign genes from another organism, rather than by the stimulus itself. By contrast, a stimulus (the sight of a predator) can generate a new belief (that there is a predator in my vicinity) in a straightforward manner. In other words, the causal chain between stimulus and response appears to be fundamentally different from that which occurs in a cognitive mental process.
Since externally generated flexibility, such as occurs in gene-swapping, differs in important respects from the learning process whereby an organism acquires new beliefs, I conclude that only if flexibility in an organism is internally generated does it require an agent-centred explanation.
I should add that if one were to envisage the acquisition of mercury resistance in mentalistic terms, one would have to view all other instances of gene-swapping between organisms in the same way. It would then follow that all organisms had minds.
F.6 Internally generated flexibility of behaviour (i.e. the ability to modify patterns of information transfer, by means of an inbuilt mechanism) is a necessary condition for the identification of cognitive mental states. Specifically, internally generated flexible behaviour in an organism is a necessary condition for the warranted ascription of cognitive mental states to it. That is, the organism must be able to modify its patterns of information transfer, by means of an inbuilt mechanism, before we can justifiably impute beliefs to it.
F.5 The occurrence in an organism of flexible behaviour does not provide a sufficient warrant for the ascription of mental states to it.
None of the biological mechanisms we have examined in this section met the requirements for internally generated flexibility. In the next section, I examine another class of phenomena which promises to deliver this kind of internal flexibility: the learning behaviour of organisms.
The following table contains a selection of definitions of the most common varieties of learning discussed in the psychological literature.
Table 2.3 - Common psychological definitions of different kinds of learning
| Kind of learning | Definition | Which organisms are capable of it? |
| Learning (general definition) |
"[A] relatively permanent change in behaviour potential as a result of experience" (Abramson, 1994, p. 2). This definition excludes changes in an organism's behaviour that are due to other factors such as physical development, aging, fatigue, sensory adaptation, or circadian rhythms (Abramson, 1994, p. 2). (I discuss this in greater detail in section 4.1 of the Appendix to chapter 2 part B.) |
Despite claims that it occurs in bacteria (Di Primio, Muller and Lengeler, 2000, p. 7), learning, in the sense used by psychologists, appears to be confined to eukaryotes (organisms such as protoctista, plants, fungi and animals, whose cells have a nucleus). Claimed occurrences of learning in bacteria appear to be actually cases of sensory adaptation, in which an organism's response to a stimulus may diminish because the organism's sensory organs no longer detect the stimulus (Abramson, 1994; Illingworth, 1999). |
| 1. Non-associative learning | "[T]hose instances where an animal's behaviour toward a stimulus changes in the absence of any apparent associated stimulus or event (such as a reward or punishment)"(Encyclopedia Britannica, 1989). Only one kind of event (the stimulus) is involved in this kind of learning. In the case of non-associative learning, "[t]he animal does not learn to do anything new or better; rather, the innate response to a situation or a particular stimulus is modified" (Abramson, 1994, p. 37). |
Found in eukaryotes (organisms whose cells have a nucleus), but not in bacteria or archaea |
| 1(a) Habituation | The decline of a response "as a result of repeated stimulation" (Abramson, 1994, p. 106). Di Primio, Muller and Lengeler (2000, p. 7) use the same definition. Example: a loud noise nearby makes you jump when you first hear it. But if you keep hearing a loud noise at regular intervals after that, you get used to it and learn to "tune it out". |
Eukaryotes (organisms whose cells contain a nucleus), including protoctista (especially protozoa, such as paramecia and amoebae), plants (Mimosa) and animals (Abramson, 1994, pp. 106, 112, 116, 117; Abramson et al., 2002). |
| 1(b) Sensitization | "[T]he opposite of habituation and refers to an increase in frequency or probability of a response" to a stimulus (Abramson, 1994, p. 105).
Example: after hearing a loud crash, smaller noises can startle a person which would otherwise go unnoticed. Non-associative sensitization occurs when an organism is exposed to an intense stimulus that elicits a strong innate response (e.g. electric shock). [N.B. Associative sensitization, known as conditioned sensitization or alpha conditioning, is defined as an increase in the probability of an innate reaction based on the pairing of two stimuli. In alpha conditioning, unlike classical conditioning, the conditioned responses arise from an innate connection. There is some debate in the literature regarding the relationship between alpha and classical conditioning (Abramson, 1994, pp. 105-106).] |
Sensitization and habituation are "ubiquitous throughout the animal kingdom. They even appear, for instance, in experiments in which the 'animals' consist of single cells or isolated ganglia" (Abramson, 1994, p. 106). |
| 1(c) Dishabituation | A "facilitation of a decremented or habituated response" (Rose and Rankin, 2001, p. 63). Dishabituation was formerly thought to be a special form of sensitization, but research on molluscs and leeches has now confirmed that it is a biologically and behaviourally distinct process (Rose and Rankin, 2001, p. 63). |
Found in most animals. |
| 2. Associative learning | A form of behaviour modification involving the association of two or more events, such as between two stimuli, or between a stimulus and a response. In associative learning, an animal does learn to do something new or better (1994, p. 38, italics mine). | All worms (with the possible exception of flatworms), and "higher" animals, such as arthropods (e.g. insects), molluscs, echinoderms and chordates (e.g. vertebrates). |
| 2(a) Classical conditioning | Classical conditioning refers to the modification of behavior in which an originally neutral stimulus - known as a conditioned stimulus (CS) - is paired with a second stimulus that elicits a particular response - known as the unconditioned stimulus (US). The response which the US elicits is known as the unconditioned response (UR). An organism exposed to repeated pairings of the CS and the US will often respond to the originally neutral stimulus as it did to the US (Abramson, 1994, p. 39). Example: a dog normally salivates (UR) at the smell of food (US). But if the dog hears the sound of a bell (CS) just before smelling the food, it will learn to associate the sound of the bell with being fed, and salivate when it hears the bell (CR). It should be noted that if the CS and US occur simultaneously, or if the CS occurs after the US, virtually no conditioning will occur. The CS needs to precede the US and be predictive of it. An animal obtains no biological advantage in learning an association between a CS and a US unless the CS can be used to predict the US. |
All worms (with the possible exception of flatworms), and "higher" animals, such as arthropods (e.g. insects), molluscs, echinoderms and chordates (e.g. vertebrates). |
| 2(b) Instrumental conditioning | Instrumental and operant conditioning are "examples of associative learning in which the behavior of the animal is controlled by the consequences of its actions... [Whereas] classical conditioning describes how animals make associations between stimuli, ... instrumental and operant conditioning describe how animals associate stimuli with their own motor actions ... Animals learn new behaviours in order to obtain or avoid some stimulus (reinforcement)" (Abramson, 1994, p. 151).
Example: a rat placed in a box with a lever presses the lever by accident, or during exploratory behaviour, and receives a small pellet of food. If this happens a few times, the rat will start to press the lever more and more often. Note: for many psychologists, the terms "instrumental conditioning" and "operant conditioning" are inter-changeable. For others, operant conditioning refers to a more sophisticated ability: an animal's "ability to operate some device - and know how to use it, that is, make an arbitrary response to obtain reinforcement" (Abramson, 1994, p. 151, italics mine). |
Defined broadly as "behaviour controlled by its consequences", instrumental conditioning is present in all animal groups (Abramson, 1994, p. 151). |
The first thing that needs to be noted in the above table is that the word "learning" is used by psychologists in a sense very different from that of ordinary usage. In popular parlance, the principal meaning of "learn", as defined by The Merriam-Webster On-line Dictionary (2004), is:
to gain knowledge or understanding of or skill in by study, instruction, or experience.
The question of which, if any, of the forms of "learning" described in the above table deserve to be called learning in the popular sense of the word, will be discussed below. For the time being, I shall resort to quote marks to indicate the psychological usage of the term.
The two main forms of non-associative "learning"are habituation and sensitization. Habituation has obvious survival value: it enables an organism to ignore stimuli that do not transmit any biologically relevant information (e.g. about food or the presence of a predator), thereby minimizing energy that may be wasted on fleeing from "false alarms" and giving it more time to engage in behaviour that enhances reproduction. Habituation also makes it possible to engage in consummatory behaviour in unpalatable but otherwise suitable situations (Abramson, 1994, p. 107).
Sensitization allows an organism to remain responsive to significant or dangerous stimuli, and to rapidly learn adaptive behaviour patterns (Abramson, 1994, p. 107).
It has been claimed that habituation occurs in all cellular organisms, even bacteria (Di Primio, Muller and Lengeler, 2000, p. 7). The evidence cited in favour of this sweeping claim (discussed at further length in section 4.2 (case study 1) of the Appendix to chapter 2 part B is the observation that following prolonged exposure to an attractant, bacteria change from a "run" to a "tumble" movement. However, if we examine the chemical basis for so-called "habituation" in bacteria, it appears to be a case of sensory adaptation - a phenomenon in which an organism's response to a stimulus may diminish because the organism's sensory organs no longer detect it - rather than habituation, where the response to a stimulus wanes because the experience of it is repeated over and over again. As Illingworth puts it:
With increasing attractant concentrations the MCPs [methyl-accepting chemically sensitive proteins on the surface of bacterial cells - V.J.T.] are progressively converted into the fully methylated state with a low affinity for the attractants (1999).
In other words, saturation of the MCPs in bacterial cells with methyl radicals reduces their sensitivity to attractant molecules. Additionally, the observation that the removal of the attractant, followed by its re-presentation after an interval of time, causes a bacterium to respond in the same way as it usually does when first exposed to an attractant, is best explained by the hypothesis that the bacterium's sensory adaptation has worn off by then. If the bacteria had been habituated, one would expect them to continue to display a diminished response (Abramson, 1994, pp. 108-109; Illingworth, 1999).
Even if one does not regard habituation as a true form of learning, there is a philosophically and biologically significant difference between failing to detect a stimulus and ignoring it: the latter may be considered as a self-initiated teleological activity on the organism's part which enables it to screen out irrelevant stimuli, the former cannot be construed as an activity of any sort, let alone one which benefits the organism.
On the other hand, certain kinds of protoctista (especially protozoa, such as paramecia and amoebae) are recognised as being capable of being properly habituated (Abramson, 1994, pp. 106, 112, 116, 117). The evidence that protoctista are indeed capable of undergoing habituation is presented and described in section 4.2 (case study 2) of the Appendix to chapter 2 part B.

The phenomenon of habituation is well-attested in Mimosa, a small shrub whose leaves are sensitive to stimulation (Abramson et al., 2002, pp. 174-176). Habituation is also known to occur in other plants: the carnivorous plant Drosera, known as the Sundew, and the Passion Flower Passionflora gracilis (Abramson et al., 2002, p. 175). I discuss habituation in plants at further length in section 4.2 (case study 3) of the Appendix to chapter 2 part B - including a possible case of associative habituation, detected in an experiment by Holmes and Gruenberg in 1965, which showed that Mimosa could discriminate between different types of stimuli: it could be "trained" to stop closing in response to water droplets, but still retained its response to the touch of a finger.
L.1 Habituation and sensitization appear to be confined to eukaryotes, or organisms with a nucleus in their cells.
In what follows, I examine the behavioural pre-requisites that an entity must satisfy before it can be credited with learning in the popular sense.
Reproducible memory
Kilian and Muller(2001) argue that any organism capable of learning should not only be able to change their behaviour, but be able to reproduce the behavioural change when the occasion demands it:
In ethology learning is defined as a change in the individual behavior which leads to a better adaptation and which is influenced by amplification and experience. Proper learning shows a reproducible learning effect... [E]pigenetic learning is based on the ability to form the interplay between stimulus input, information transfer, memory, and behaviour in an individual and reproducible way (2001, pp. 1 - 2, italics mine).
If I read Kilian and Muller correctly, the insight underlying their argument is that learning is a teleological process, which benefits its possessor. On strictly logical grounds, the benefits of learning - better adaptation - can only accrue if what is learned is remembered, in the minimal sense defined above: the individual must be able to store new information in response to changing circumstances. Kilian and Muller's stipulation of a "reproducible learning effect" is not a logical but a methodological criterion that an organism must satisfy before scientists can justifiably attribute learning to it.
On the other hand, the possession of memory by an individual does not guarantee its ability to learn. An entity's ability to store new information about its surroundings does not imply that it can acquire new skills, let alone knowledge or understanding. I shall return to the notion of a skill below, in the section on associative learning.
L.3 Learning should not be attributed to an organism unless it displays a change in its pattern of behaviour which it is able to reproduce on a subsequent occasion.
L.2 The existence of memory in an organism is a necessary but not a sufficient condition for learning.
Flexible behaviour
An additional requirement for "true" learning is flexibility of response patterns. According to the popular definition cited above, an individual that learns, thereby gains something - knowledge, understanding or a skill - from its experience. But it is logically impossible for an organism to manifest its new knowledge, understanding or skills if its behaviour patterns are fixed, according to the definition given above. We may therefore formulate the following conclusion:
L.4 The ability of an organism to display flexible behaviour is a necessary condition for learning.
The foregoing conclusion entails that organisms undergoing habituation are not learning to respond in a new way, but are simply diminishing their innate response to a stimulus, after repeated exposure to it. The waning in their response over the course of time does not represent a change in their pattern of behaviour, but can be described as a continuance of an existing fixed pattern. Putting it mathematically, we can describe an organism's response to a stimulus in terms of a function F, whose inputs include not only the intensity of the stimulus but also the number of exposures to it. During habituation, the function F does not change; all that changes over time is the value of one of the parameters (number of exposures). This is an historical parameter, but as we have seen, nothing in the definition of "fixed pattern" given above prohibits us from using time-dependent variables to describe animal behaviour. If the behaviour of an organism undergoing habituation conforms to a fixed pattern, then it provides us with no grounds for imputing mental states to it (Conclusion F.2). The same remarks apply to sensitization, which is the opposite of habituation (Abramson, 1994, p. 105).
Although psychologists customarily refer to habituation as a kind of "learning", their definition of learning (a relatively permanent modification in an organism's behaviour as a result of experience) overlooks the possibility of using historical parameters (such as number of exposures) as inputs to behavioural functions; instead, historical changes are simply treated as generating a new behavioural function.
L.5 The ability of an organism to undergo (non-associative) habituation and sensitization is not a sufficient condition for learning, in the popular sense of the word.
L.6 The occurrence of non-associative habituation and sensitization in an organism does not provide a sufficient warrant for the ascription of mental states to it. (Corollary of Conclusion F.2.)
We may conclude, then, that habituation does not necessarily constitute true learning, and that senses, memory, indirect stimulus-response coupling and habituation are not, by themselves, sufficient to establish the existence of cognitive mental processes in an organism (see Conclusions S.6, M.2, F.2 and L.4).
It should be stressed that habituation is a phenomenon which admits of varying degrees of complexity in the animal world. The foregoing conclusions apply only to the simplest cases, where the change in behaviour is a non-associative one. In some animals, a variety of circumstances can cause the response to the original stimulus, attenuated by habituation, to re-appear: a change in some "dimension" of the stimulus (e.g. a change in the pitch or volume of a sound); the passage of time; the presentation of a new stimulus, like the original one (dishabituation); a new context for the stimulus; and fatigue (Balkenius, 2000). Balkenius (2000) regards habituation as a process where an organism learns what to expect in a certain situation or context. Context-dependent learning will be discussed later; what we are considering here is purely non-associative "learning".
We have seen that the definition of learning used by psychologists is broader than the sense captured by popular usage. However, Kilian and Muller (2001) employ a more restrictive definition of learning, in which "an animal does learn to do something new or better" (Abramson, 1994, p. 38). This corresponds to what psychologists call associative learning.
Kilian and Muller (2001, pp. 3 - 4) contrast the non-associative forms of behaviour modification found in unicellular organisms with that of multicellular organisms, where the cells involved in chemical information transfer retain their functional specificity, but this specificity is de-coupled from the "goal" of the behaviour. At the beginning of the individual's life, the goal is unspecified or "open-ended": the synaptic connections are not fixed. The same stimulus may be linked to any one of a multitude of responses. Different paths open up, but the final selection from the range of goal cells that come into play is made by the stimulus in the individual's local external environment (e.g. in imprinting, the first object the individual happens to see), leading to an individualised, reproducible learning effect. Paths between sensors and effectors vary from one individual to another, and are not genetically determined.
I would suggest that Kilian and Muller's claim, that functional specificity is retained at the chemical (micro) level but that there is no goal specificity at the macro level, can be better understood if we imagine that:
(i) each kind of behaviour exhibited by an organism can be described by a higher-level function (or combination of functions);(ii) higher-level functions invoke various lower-level functions within the organism which describe its interactions at the chemical level;
(iii) although the higher-level functions that describe an organism's response to various stimuli are not fixed at the beginning of its life, the lower-level functions they invoke constitute a fixed, finite set. Each higher-level function invokes a succession of several functions from this fixed set. A simple example: z = F(x) = f(g(h(x))), where: x is an input variable corresponding to the intensity of a certain kind of stimulus; f, g and h are three low-level "chemical" functions that operate successively on the same stimulus; F is a higher-level function that describes a certain kind of behaviour; and z is the behavioural output corresponding to F;
(iv) the organism possesses an internal mechanism allowing it to modify its higher-level functions. In the above example, the definition of F(x) might change by invoking another of the oranism's fixed repertoire of low-level functions (e.g. F(x) = f(g(j(x))) instead of f(g(h(x)))) or invoking the functions in a different order (e.g. F(x) = f(h(g(x))), instead of f(g(h(x)))).
Drawing on the work of Dretske and Dennett, Beisecker (1999, p. 298) proposes a slightly more restrictive definition of learning: organisms are capable of learning if they have the "ability to tailor their own responsive dispositions to their particular surroundings". On this interpretation, even imprinting would not qualify as true learning, as the behavioural response, once formed, cannot be subsequently modified to fit different circumstances (e.g. the death of the "parent" imprinted on the newborn individual's memory).
It remains to discuss whether associative learning is a sufficient condition for the occurrence of learning in the popular sense of the word, and whether it indicates the occurrence of mental states in an organism.
Abramson (1994, p. 38) defines associative learning as:
a form of behaviour modification involving the association of two or more events, such as between two stimuli, or between a stimulus and a response. In associative learning, an animal does learn to do something new or better (1994, p. 38, italics mine).
There are two broad categories of associative learning: classical conditioning and instrumental or operant conditioning. These terms are defined in Table 2.3 above. Pavlov's experiment, in which he trained a dog to salivate at the sound of a bell (which it associated with being fed) is an example of classical conditioning. A rat that learns to associate one of its bodily actions (pressing a lever) with a desired result (obtaining food) is an example of instrumental conditioning.
Both kinds of associative learning are biologically beneficial to their possessors. According to Brembs (2000, p. 2), classical conditioning enables organisms in the wild to associate biologically neutral stimuli with significant ones, enabling them to make better predictions about their environment, while operant conditioning reinforces behaviour that satisfies their appetites or enables them to avoid aversive stimuli.
The examples of classical and instrumental conditioning just given related to animals (dogs and rats) whose possession of mental states is almost universally acknowledged. The question we have to address, however, is whether we should ascribe mental states such as beliefs and desires to any animal that is capable of undergoing conditioning - a worm, for instance.
The significance of associative learning with regard to agency
The significance of associative learning from the standpoint of intentional agency is that it allows an individual to associate acts with consequences. In the absence of such an association, it is difficult to envisage how agency could occur at all.
Associative learning qualifies as flexible behaviour
Associative learning certainly qualifies as flexible behaviour according to the definition we have given. It is not fixed, as the value of the output variable (i.e. the response) for the same input variable (stimulus) does not remain the same over time. There is genuine novelty here, which cannot be treated as a temporal extension of an existing pattern of activity within the organism by introducing extra historical variables, as we did with habituation. Instead, what we see here are either new conditions for activating an existing behaviour pattern (classical conditioning), or the emergence of a new behaviour pattern (instrumental or operant conditioning). In a simple case of classical conditioning, the organism learns to respond to a new stimulus (the conditioned stimulus) in the same way as it does to an existing one (the unconditioned stimulus). This is flexible behaviour, because one of the programs governing an organism's behaviour changes over time: there is a change in the conditions under which one of its behaviour function(s) is activated. In operant conditioning, the organism acquires a new behavioural function through "trial-and-error learning". Once again, this requires a program change.
Associative learning qualifies as internally generated flexible behaviour
As we saw in our discussion of bacterial gene-swapping in the section on flexible behaviour, the program governing a bacterium's response to mercury does not modify itself: it receives new, pre-packaged instructions from an external source (another bacterium). While this qualifies as flexible behaviour, it differs in important respects from what Kilian and Muller (2001) would characterise as true learning. Lack of goal specificity is a vital feature of their account: the same stimulus may be linked to any one of a multitude of responses. Moreover, it is the stimulus in the individual's local external environment which determines the specific response that is displayed by an individual organism, whereas in gene-swapping, as we saw above, the change in response bears no relation to the stimulus itself.
L.7 The occurrence in an organism of flexible behaviour is not a sufficient condition for learning.
In the case of associative learning, however, the new behaviour is acquired through an internal learning mechanism. This in-built mechanism for acquiring information allows the individual to modify its response to a stimulus.
L.8 The capacity for associative learning in an organism is a sufficient condition for its being able to engage in internally generated flexible behaviour.
It has been claimed that paramecia possess a capacity for learning through classical conditioning. The original experiment was reported in a study by Hennessey, Rucker and McDiarmid (1979) and is still widely quoted (e.g. by Martin and Gordon, 2001). I discuss this study in section 4.3 (case study 1) of the Appendix to chapter 2 part B and conclude that since no-one has yet relicated or even attempted to replicate the results, it would be unwise to rely on them at this stage. Also, further research is needed to explain why experiments (of a different kind) on another kind of ciliate (Stentor) failed to demonstrate instrumental conditioning (Wood, personal email, 18 June 2003; Hinkle and Wood, 1994).
Research into the possibility of classical conditioning in mimosa plants has produced negative or conflicting results, and the methodology of studies which found conditioning has been criticised (Abramson et al., 2002, pp. 175-176). In keeping with my methodological constraints, these studies will be ignored here. Section 4.3 (case study 2) of the Appendix to chapter 2 part B contains a summary of research conducted to date, including a detailed discussion of a more refined plant learning experiment by Abramson et al. (2002, pp. 173 - 185) on Philodendron cordatum plants, which yielded negative results. As there are several possible explanations for this failure, follow-up research with longer intervals is recommended by the authors.
Although protoctista and plants are highly adaptive, multicellular, eukaryotic organisms which are capable of being habituated (unlike bacteria), there is no good evidence to date that they are capable of associative learning. Further research into the alleged learning abilities of these organisms is needed to settle the matter, but in the light of what we know, it seems prudent to regard them as lacking beliefs and desires.
The relationships between the different groups (or phyla) of worms, and other animals, are discussed in further detail in section 4.3 (case study 3) of the Appendix to chapter 2 part B. Of special interest here are flatworms (believed to be the most "primitive" group) and roundworms (or nematodes), the group to which the well-studied Caenorhabditis elegans (usually abbreviated as C. elegans) belongs.
Even the simplest flatworms have been credited with the capacity for associative learning. However, the attribution of associative learning to even the simplest flatworms is controversial, as different authorities use definitions of "associative learning" and some of the effects of associative learning can be mimicked by phenomena that can be explained in non-mentalistic terms. It is no easy matter for scientists to verify that an animal has undergone classical conditioning and/or instrumental conditioning (two common forms of associative learning which are defined in the table above), as these processes are easily confused with other behavioural processes in animals which do not involve learning - for example, pseudo-conditioning and sensitization. The problems involved in verifying scientifically that an animal has undergone classical and/or instrumental conditioning, and the history of previous mis-identifications of conditioning in worms, are discussed in section 4.3 (case study 3(a)) of the Appendix to chapter 2 part B.
I have not been able to locate any studies to date showing unequivocally that flatworms (platyhelminthes) are capable of classical conditioning. However, recent research on another worm, the well-studied Caenorhabditis elegans, has demonstrated that even worms with very "simple" nervous systems are capable of associative learning - in particular, classical conditioning. C. elegans belongs to the phylum Nematoda (roundworms) and is a favourite of scientists studying the genetic and molecular bases of learning, because it has a fully mapped nervous system with only 302 neurons and a small, almost completely sequenced genome. Although roundworms, like flatworms, are protostomes (animals with one opening that serves both as a mouth and an anus), they are not closely related to flatworms.
The evidence that C.elegans is truly capable of undergoing classical conditioning is discussed in section 4.3 (case study 3(b)) of the Appendix to chapter 2 part B. In particular, recent studies have shown that C. elegans worms can actually be conditioned to radically alter their preferences: they will avoid a stimulus they had formerly been attracted to, after it has been paired with an aversive stimulus. This change of preference cannot be explained away as "conditioned sensitization" because the old response is not re-awakened. The worms are actually learning to do something new: they are changing their pattern of response to a stimulus.
Additionally, Catharine Rankin, who specialises in learning mechanisms in C. elegans, claims (personal email, 31 May 2003) that recent work has established that it can indeed associate a CS with a US.
Why should a central nervous system be essential for learning?
Kilian and Muller argue that because unicellular organisms have "genetically fixed ... networks of information transfer" (2001, p. 3), they are unable to modify their own information transfer pathways and so cannot learn to respond to a new stimulus:
Unicellulars are not able to synthesize or modify cellular substances not already coded genetically in a fixed way. So, they cannot install new, individualized, ... reproducible information transfer paths within their life-time... The synthesis of new substances for new irreversible and reproducible information transfer paths as an answer to a new, formerly not identifiable stimulus is not possible in unicellulars. Therefore, unicellulars seem most certainly not able to learn, at least according to an ethological definition (2001, pp. 2-3, italics mine).
As we have seen, the phenomenon of gene-swapping, a flexible form of behaviour which is found even in unicellular organisms, differs in significant ways from learning. Gene-swapping is not a process whereby an organism modifies its own information transfer paths in response to a stimulus, but rather a process whereby the organism's information transfer paths are externally modified by processes unrelated to any stimulus. More controversially, Kilian and Muller propose that the kind of information transfer functionality required for true learning is possible "only when several cells of information transfer functionality come into close contact spatially, i.e. in organisms with central nervous systems" (2001, p. 4, italics mine).
It remains to be seen whether Kilian and Muller's theoretical argument for confining learning to organisms with central nervous systems is a valid one, but the empirical evidence in support of such a conclusions appear to be reasonably well-established:
L.9 Associative learning appears to be confined to organisms with central nervous systems. It is found in most but possibly not all phyla of animals with central nervous systems. (Flatworms may not be capable of associative learning, but many other phyla of worms are.)
Or more generally:
F.7 Internally generated flexible behaviour appears to be confined to organisms with central nervous systems. It is found in most but possibly not all phyla of animals with central nervous systems. (Flatworms may not be capable of it, but many other phyla of worms are.)
Combining this result with conclusion F.6 yields the following biological criterion for intentional agency:
B.6 We can only identify cognitive mental states in organisms with central nervous systems.
Evidence for instrumental and operant conditioning
There are two major methodological concerns in relation to instrumental conditioning. First, it is easily confused with classical conditioning. Some examples are given in section 4.3 (case study 3(c)) of the Appendix to chapter 2 part B. Nonetheless, it is generally acknowledged that instrumental conditioning - defined broadly as behaviour controlled by its consequences - occurs in all animal groups (Abramson, 1994, p. 151).
The other major methodological worry regarding instrumental and operant conditioning is the lack of an agreed definition for operant conditioning. Many psychologists use the terms "instrumental conditioning" and "operant conditioning" (or operant learning) interchangeably to mean something like "behaviour controlled by its consequences" (Abramson, 1994, p. 151). A few researchers follow Skinner in reserving the term "operant learning" for behaviour which "appears more flexible and adaptable" (Grau, 2002, p. 85) than instrumental conditioning. Abramson (1994) also makes a distinction between "instrumental conditioning", and "operant behaviour", reserving the latter term for special cases. He remarks that if we adopt the common definition of operant behaviour, then
...such behavior is present in all animal groups. However, if operant behavior is defined in terms of its functional influence on the environment and the ability to use an arbitrary response, then... [it] is limited to vertebrates and perhaps some species of mollusks, crustaceans, and insects. A rule of thumb I have found useful... is that in operant behavior, an animal must demonstrate the ability to operate some device - and know how to use it, that is, make an arbitrary response to obtain reinforcement. (1994, p. 151, italics mine).
However, Abramson's distinction between instrumental and operant conditioning has been queried by other researchers - for instance, Bjorn Brembs considers Abramson's definition to be "more obfuscating than enlightening" (personal email, 11 August 2003):
What does he mean by "device"? How does he test that the animal "knows"? (personal email, 11 August 2003).
I shall revisit these criticisms in section 2.B.5, where I propose a more stringent definition of operant conditioning.
In any case, what is not disputed is that simple worms such as C. elegans are capable of undergoing instrumental conditioning. Recent experimental evidence for what may be a case of operant conditioning in C. elegans is presented in section 4.3 (case study 3(d)) of the Appendix to chapter 2 part B. I advise caution in the interpretation of these experiments, as the studies performed produced some conflicting results, for reasons that are not altogether clear.
After surveying the literature and examining the available evidence, I conclude that many (probably most) phyla of worms and all "higher" animals are capable (to some degree) of undergoing two forms of associative learning - classical and instrumental conditioning. That leaves two philosophical questions unanswered. First, is this learning in the standard (as opposed to psychological) sense of the word? Second, do we need to explain it in terms of mental states?

Earlier in this chapter, I suggested that the word "learning" be reserved for mental states, out of respect for norms of popular usage. However, it is not absolutely clear that popular usage dictates that learning is a mental state: according to the dictionary definition cited above, acquiring a skill by experience also qualifies as learning. There is good evidence that some worms, whether or not they possess mental states, are capable of acquiring skills through experience. They can be conditioned to avoid a stimulus that they were previously attracted to. Since their preference-changing behaviour is internally generated, flexible, and biologically useful to them, it surely deserves to be called a skill. I therefore suggest that we should regard conditioning as a form of experience (i.e. training) without committing ourselves at this stage as to whether worms are intentional agents. Being conditioned to change one's preferences is therefore acquiring a skill through experience. It follows that some worms can learn, regardless of whether they have mental states or not. They are "educable" creatures.
L.10 The ability of an organism to undergo associative learning (classical and/or instrumental conditioning) is a sufficient condition for its being able to learn, in the ordinary sense of the word.
To sum up: recent research has shown that roundworms (nematodes) are capable of associating stimuli when they undergo classical conditioning. These simple worms represent a breakthrough in mind-like behaviour: they are educable creatures, capable of true learning, as defined by Kilian and Muller (2001). However, it has not yet been shown that they are capable of genuine trial-and-error learning (operant behaviour, as defined by Abramson).
A note on Dennett's mental hierarchy
Dennett (1997, p. 112) describes organisms with a capacity for associative learning as "Skinnerian creatures". They occupy the second level in his hierarchy of mind-like organisms (1997, pp. 109 ff.). Do roundworms qualify as "Skinnerian creatures"? If we adopt Dennett's definition of "Skinnerian creatures" as organisms with a capacity for associative learning, whose response patterns can be altered by their experiences, the answer is yes. However, Dennett also defined Skinnerian creatures as animals that are capable of trial-and-error learning from their past mistakes and successes, i.e. operant conditioning (1997, p. 112). On this criterion, it is not clear that roundworms qualify. It may turn out that there are creatures that are capable of associative learning but incapable of operant behaviour. Dennett may have conflated two stages in his mental hierarchy.
From the beginning, some pioneers of learning research (notably Pavlov) have championed a non-mentalistic account of conditioning. Although Pavlov's stimulus substitution theory of associative learning is no longer considered an adequate account (see section 4.4 Appendix to chapter 2 part B), there is still no scientific consensus regarding the cognitive requirements of associative learning.
However, some philosophers have championed a mentalistic account of conditioning - especially instrumental conditioning. Dretske (1999) considers creatures that can undergo conditioning as "educable", and credits them with beliefs that are acquired and modified by the associations they form. Beisecker (1999) defines "educable" creatures somewhat differently, as organisms that have expectations that they can bring about certain outcomes. These creatures continually revise their expectations by being sensitive to the consequences of their actions. Although both accounts acknowledge that educable animals can make mistakes, Beisecker's account (which I discuss in section 2.B.7) lays special emphasis on the ability of animals to adjust their expectations.
Dretske: Why associative learning is evidence for belief and agency in animals
Dretske (1999) has argued that animals which can undergo operant conditioning are capable of not only memory and learning, but also belief and agency. He carefully differentiates agency from behaviour: behaviour may have a meaning, but purposeful acts are governed by their meaning. When an action occurs, events that have a meaning cause an animal to behave in a certain way, by virtue of their meaning. With computers and plants, on the other hand, events which have a meaning we can recognise, cause something to behave, but it is the intrinsic properties of the events, and not their meaning, that explains the behaviour.
Dretske illustrates his point with a few well-chosen examples. A speaker utters the words "Vibrate rapidly" into a microphone. The microphone's diaphragm vibrates rapidly. The sounds made by the speaker had a meaning - "Vibrate rapidly" - but it is not what the speaker says, but the physical properties of the sounds, which cause the diaphragm to vibrate rapidly. (The microphone would have vibrated rapidly even if the speaker had said, "Be still".) The microphone is sensitive to sound, not meaning.
A thermostat turns the heat on and off, keeping the room at a comfortable temperature. Is this mere behaviour or is it action? We could say that the thermostat simply behaves because it lacks beliefs and desires: a goal-centred intentional stance suffices to explain its behaviour. But Dretske goes beyond this obvious response and attempts to explain what would be needed to make the thermostat's behaviour a bona fide action.
Most thermostats have a bimetallic strip that functions as a thermometer (its degree of curvature represents room temperature) and as a switch - if the room cools to the desired temperature, the strip touches an adjustable contact, whose position corresponds to the room's desired temperature, thereby closing an electrical circuit to the furnace and turning the heat on. The thermostat senses a drop in temperature and corrects it. Although the curvature of the thermostat's bimetallic strip means something (it represents the ambient temperature), it is not the meaning, but the curvature, that governs the thermostat's behaviour. Dretske argues that if we take away the meaning but keep the curvature (e.g. by bending the strip with a pair of pliers), the thermostat will behave the same as it would if the room were cool.


Now consider a foraging bird, who tries to eat a Monarch butterfly that has been reared on a toxic form of milkweed. Eating the butterfly makes the bird vomit. The next day, the bird sees a Viceroy butterfly, which looks remarkably like the poisonous Monarch. The bird flies away. Why does the bird not eat the Viceroy? Its behaviour can readily be explained in terms of operant conditioning: it learned to avoid the stimulus (Monarch butterflies) after a punishing experience, and its internal representation of a Monarch butterfly caused it to avoid a similar-looking butterfly. Whereas a thermostat is hard-wired to behave as it does, the bird's behaviour is triggered by its memory (stored internal representation) of an unpleasant experience, where it learned that a Monarch was poisonous.
Learning of this sort (operant conditioning) consists in harnessing these internal representations to control circuits so that behaviour will occur in the external conditions on which its success depends. Like the thermostat..., this internal representation ... has both a meaning and a causal role, but, unlike the instrument..., its meaning explains its causal role" (Dretske, 1999, p. 10, italics mine).
As Dretske makes no distinction between instrumental and operant conditioning, I shall assume that he uses the two terms interchangeably (unlike Abramson, 1994, who distinguishes between them).
The nub of Dretske's account is that many animals have a learning history which imparts a meaning to their experiences. It is this meaning which explains their causal role in animal behaviour. When animals use what they have learned to achieve their goals, they can be described as agents. Of course, biologically adaptive hard-wired behaviour (e.g. reflexes) in animals is not agency: it is not learned but automatic, and its efficacy is independent of any meaning (or biological purpose) it may have. Using our terminology, we might say that unlearned behaviour can be explained in terms of a mind-neutral, goal-centred intentional stance:
L.11 An organism must be capable of learning before it can be said to have cognitive mental states.
In the passage cited above, Dretske equated learning with success, it enables an organism to achieve its goals. The organism does this by drawing upon past experiences (which it stores as internal representations). These experiences can be said to associate acts (e.g. eating a Monarch butterfly) with consequences (e.g. becoming ill).
This brings us to the crux of the matter: it makes no sense to ascribe agency to an individual unless it is capable of learning the consequences of at least some of its actions. The significance of associative learning is that it appears to be the most fundamental way in which organisms (on Earth, at least) connect acts with consequences. Associative learning is thus the most fundamental form of true learning.
L.12 An organism must be capable of associative learning before it can be said to have cognitive mental states.
The bird's internal representation causes the avoidance behaviour precisely because it means something about its external environment (i.e. that a certain kind of butterfly is present - the sort of butterfly the bird, after its experience of vomiting, wants to avoid). In this case, we cannot isolate the meaning from the physical properties of the representation and say that the latter, and not the former, cause the behaviour. Here, according to Dretske, we have bona fide agency: the bird has learned the Monarch butterflies are poisonous, it believes that the Viceroy butterfly is a Monarch, and it flies away because it wants to avoid the Viceroy. Dretske uses the word "belief" rather than "knowledge" here, because the bird is in fact mistaken.

Plants, on the other hand, cannot act, according to Dretske. The adaptive behaviour of the Scarlet Gillia illustrates why. Even though this plant can change colour from red to white in summer, thereby attracting pollinators, its adaptive behaviour is triggered not by its "meaning" or biological significance, but by the intrinsic properties of the chemical switches that cause its behaviour, coupled with the historical fact that similar behaviour by its evolutionary forebears, from whom it inherited its genes, enables it to reproduce successfully.
In my opinion, Dretske does an excellent job of explaining why we say that machines and plants behave rather than act, but his defence of agency in animals is marred by an equivocation between two senses of the word "meaning" - one of which can be characterised in terms of a goal-centred intentional stance ("significance" or "purpose"), while the other requires an agent-centred stance ("aim" or "intention"). Dretske makes no attempt to differentiate between them. A thermostat's behaviour has a "meaning" for us as agents: we set it at a level that makes us feel comfortable. On the other hand, there is no need to ascribe intentions to the Scarlet Gillia to explain its behaviour: its "meaning" can be characterised using a goal-centred stance, as it is biologically adaptive. What about conditioned behaviour in animals?
Dretske has made it clear (1995) that he regards animals with beliefs as agents. If Dretske were trying to rigorously demonstrate that conditioned behaviour in animals could be described as agency simply because it could be characterised in agent-centred terminology, then he would indeed be begging the question. But in fact, Dretske seems to be making a suasive case (rather than a strictly logical one) for belief in animals that undergo instrumental or operant conditioning. He appears to be arguing that a belief-based account makes sense of the fact that the bird changes its behaviour because of what it remembers:
Notice how natural it is in this case (unlike the case of the plant or the thermostat) to explain the bird's behavior in terms of what it believes. It is natural, I submit, because memory about some previously experienced object is so obviously implicated in why the bird behaves as it does... Talk of memory becomes appropriate here because behavior changes after a perceptual encounter.. The thermostat and the plant come into the world hard-wired to behave the way they do. What happens to them is not relevant to why they behave that way... Not so with the bird (1999, pp. 28-29, italics mine).
If we look at Dretske's writings, we can discern two criteria by which he claims to distinguish between organisms with and without beliefs. The wording above suggests the following criterion:
Dretske Mark I:
We can use the criterion of learned, flexible behaviour to distinguish between believing and non-believing organisms: whereas the thermostat and the plant are "hard-wired" (1999, p. 29) to behave as they do, the bird is not. It can learn.
This criterion is a little muddled: as we saw above (Conclusion F.4), flexible behaviour is a universal feature of organisms, including plants. Dretske employs the terms "mindless", "unlearned" and "hard-wired" inter-changeably. However, learned flexible behaviour is restricted to certain kinds of animals. Can we draw the line here?
Dretske's case would collapse at once if a parallel case could be found where he would be unwilling to ascribe belief to the organisms concerned. I discuss three such alleged cases in section 4.5 of the Appendix to chapter 2 part B: conditioning of the autonomic nervous system, of leg withdrawal in headless cockroaches, and of flexion of the hind legs within the spinal cords of paralysed rats.
These three examples undermine the notion that flexible behaviour patterns - even those that are generated by an internal mechanism (Conclusion F.6) - can serve to distinguish organisms with beliefs from those that lack them.
Gould (2002) has argued that the process whereby animals learn to form associations is too innate to qualify as genuine cognition:
To most minds ... cognition implies an ability to step outside the bounds of the innate, including the innate wiring that enables animals to learn through classical and operant conditioning. It means, instead, a capacity to perform mental operations or transformations and thus to plan or make decisions (Gould, 2002, p. 41, italics mine).
Gould's argument appears to confuse learning (which is not innate) with its mechanism, which is necessarily innate, as Tooby and Cosmides (1997) argue:
To learn, there must be some mechanism that causes this to occur. Since learning cannot occur in the absence of a mechanism that causes it, the mechanism that causes it must itself be unlearned - must be "innate" (1997, p. 18, italics mine).Gould's case is further weakened by his damaging admission (2002, p. 44) that even human cognition may not be flexible enough to meet his exacting criteria - as well as his use of obscure terminology ("mental operations", "decisions") to elucidate the meaning of "cognition". However, his negative point stands: there is no need to explain behaviour patterns whose flexibility is internally generated, in mentalistic terms.
We can make this point more clearly if we think of the new behavioural patterns acquired by organisms as skills that enable it to survive. All organisms have survival skills, by virtue of their history. What is unusual about learning organisms is that they have an internal mechanism that allows them to vary their patterns of responding to their surroundings by acquiring new survival skills. But the acquisition of a skill need not be envisaged in mentalistic terms: a mind-neutral, goal-centred stance appears adequate to characterise it.
F.8 The presence in an organism of flexible behaviour patterns that are acquired through an internal mechanism does not provide a sufficient warrant for our being able to ascribe cognitive mental states to it.
As we have already seen, the acquisition of a skill through experience constitutes the simplest kind of learning, if we use the word "learning" in its ordinary sense. It follows that learning, in its simplest form, can sometimes be understood in mind-neutral terminology. Following the methodology I have adopted, we should prefer this mind-neutral interpretation unless a mentally richer account proves to be more scientifically productive.
As we saw in section 2.B.4(c), associative learning can be viewed as the acquisition of a skill through experience. We can now formulate the following two conclusions regarding learning-related criteria for identifying organisms' mental states:
L.14 A capacity for associative learning in an organism does not provide a sufficient warrant for our being able to ascribe cognitive mental states to it.
L.13 A capacity for learning in an organism does not provide a sufficient warrant for our being able to ascribe cognitive mental states to it.
All learning, including human language acquisition, is to some extent innately driven, as Gould himself acknowledges (2002, p. 44). But instead of pessimistically concluding that "by the strictest standards, perhaps there is no genuine cognition in any species, our own included" (2002, p. 44), I believe it would be more profitable to search for cognition by examining what it is that newly acquired behaviour patterns enable their owners to do.
Dretske's second criterion for distinguishing organisms with beliefs from those without (which I shall call "Dretske Mark II"), addresses this very question. I shall return to it in section 6, when I discuss representations and mental states, after my discussion of how controlled movement in organisms can help us to identify their mental states.
To sum up: while associative learning is an internally driven, flexible process, which underlies all other forms of true learning and enables an animal to acquire valuable new survival skills, it does not appear to be a sufficient condition for having beliefs. Something more is needed.

Case study: agency in viruses?

A schematic representation of the HIV virus.
Copyright Dr. Shaun Heaphy, Tulane University.
The phenomenon of phenotypic plasticity is perhaps the best prima facie candidate for agency in viruses. Phenotypic plasticity can be defined as the ability of organisms with the same genotype to vary their developmental pattern, phenotype or behaviour in response to varying environmental conditions (Ancel and Fontana, 2002).
A well-known case of phenotypic plasticity in viruses is the lysis-lysogeny decision, in which parasitic lambda-phage viruses adopt a bet-hedging strategy when they invade a host bacterium: they may either "decide" to kill the host immediately by multiplying until the host's cell walls burst (lysis) or to remain quiescent and confer immunity to infection upon its host (lysogeny). The strategy is described in detail in section 5.1 of the Appendix to chapter 2 part B.
Preuss's (2000) article describes the viral DNA as making a decision. Should we take this literally? If not, why not?
First, as the article states, it is random thermal background noise which determines whether the viral DNA is expressed or remains quiescent. It would be a misuse of the English language to describe this as a decision: decisions, by their very nature, require rational justification.
Second, the behaviour displayed by the viruses is in no way self-initiated: it is driven entirely by external environmental changes (thermal fluctuations). Before we can describe a piece of behaviour as a "decision", there has to be some kind of agency involved. At the very least, internal states of the organism (as well as external conditions) must influence the behaviour observed.
The foregoing discussion points to two conclusions regarding the kind of behaviour that can serve to identify intentional agency (and hence mental states) in organisms:
A.2 Behaviour by an organism must vary in response to its internal states, as well as external conditions, before it can be regarded as a manifestation of a cognitive mental state.
A.1 Behaviour by an organism must vary in response to non-random internal states before it can be regarded as a manifestation of a mental state.
Our ability to describe an entity's behaviour according to the intentional stance is a necessary condition for our being able to ascribe cognitive mental states to it (Conclusion I.1). The intentional stance characterises the behaviour of an organism as movement towards a goal which it has information about. For this reason, directed movement is a requirement of intentional agency:
A.3 An organism must be capable of directed bodily movements before these movements can be regarded as a manifestation of a cognitive mental state.
The term "directed movement" is not meant to suggest whole-body locomotion. The question we have to address is: is directed movement a sufficient warrant for the ascription of intentional agency to an organism?
Case study: bacteria
Common bacteria like E. coli are capable of directed movement towards or away from the objects they sense. For instance, they swim in chemical gradients towards attractants (e.g. glucose) or away from repellents (e.g. benzoate) - a phenomenon known as chemotaxis (Di Primio, Muller and Lengeler, 2000, pp. 4 - 5). Other bacteria display phototaxis and magnetotaxis, or directed movement in response to light and magnetic fields, respectively (Martin and Gordon, 2001, p. 219).
The occurrence of directed bodily movement in bacteria (and, as we shall see, protoctista and even plants), suggests the following conclusion:
A.4 All cellular organisms are capable of directed movement.
A bacterium actually has two kinds of movement: directed movement (known as a run, where the bacterium keeps swimming in the same direction) and random motion (known as a tumble, where the bacterium randomly changes direction). The mechanism is described in section 5.2 (case study 1) of the Appendix to chapter 2 part B. What it shows is that the movement of bacteria, while directed, is not in any way flexible, and hence does not warrant a mentalistic interpretation (see Conclusion F.3). This points us to a negative conclusion:
A.5 The occurrence of directed bodily movement in an organism does not provide a sufficient warrant by itself for the ascription of mental states to it.
Protoctista and plants

Are there, perhaps, more complex kinds of directed bodily movement which require a mentalistic explanation? I discuss the directed movement of protoctista and plants in section 5.2 (case studies 2 and 3) of the Appendix to chapter 2 part B, where I conclude that there does not seem to be any reason to treat the directed movements of protoctista as significantly different from the directed movement of bacteria. By contrast, the directed movement of plants is almost as varied and complex as that of animals. However, complexity per se does not warrant the ascription of mental states (see Conclusion C.3). Unless the behaviour is also flexible, we should adopt a mind-neutral intentional stance towards it (see Conclusion F.3). Nothing that I have read in the literature suggests that plant movement is flexible according to the defintion used in this thesis.
On the other hand, as Di Primio, Muller and Lengeler persuasively argue (2000, p. 10), there appears to be no inherent reason why the absence of locomotion in plants, fungi and certain animals should preclude the possibility of cognition on their part:
A.6 A capacity for local movement (locomotion) in an organism is not a requirement for its possession of mental states.
Albrecht-Buehler (2003a) believes that animal cells possess a kind of intelligence, and criticises the view that these cells are "rigidly operating chemical machines that derive their operating instructions internally from their genes and externally from chemicals and electrical signals emitted rigidly by other cells." He has made some intriguing claims regarding the centrosome, a spherical area near the nucleus of a cell, which (in animal cells but not in most plant cells) contains a pair of cylindrical structures called centrioles. Albrecht-Buehler claims that the centrosome is actually the control centre (or "brain") of an animal cell, while the centrioles function as the cell's "eyes". These "eyes" can detect objects and other cells by pulsating near-infrared signals, and steer the cell towards their source. (The movement of animal cells differs from the phototactic behaviour of bacterial cells in several significant ways: bacteria, although light-sensitive, cannot see objects.) This means that cells can order and integrate a large amount of visual data.
Albrecht-Buehler believes that animal cells can navigate (see diagram), and that this ability is a manifestation of cell intelligence. He suggests that "the best place to start searching [for cell intelligence is] the field of cell movement."
This raises the question: is navigation a necessary condition for the possession of mental states? If we define navigation broadly to include any kind of sensory guidance, the answer is in the affirmative, as both sensory capacities (Conclusion S.3) and directed bodily movement (Conclusion A.3) are pre-requisites for the ascription of mental states to organisms. As Albrecht-Buehler puts it:
A moving cell has to operate its own body in sophisticated ways and, in addition, may have to navigate in space and time while dealing with numerous unforeseeable events, such as encounters with other cells and other objects that its genome could not possibly have anticipated. I think that cell motility, indeed, revealed cell intelligence.
We can express ourselves more rigorously as follows:
A.7 An organism must be capable of navigation before its movements can be regarded as a manifestation of a cognitive mental state.
Broad definition - "navigation"
Any organism that can use its senses to steer itself or a part of its body around its environment is capable of navigation.
I have added the phrase "or a part of its body" to the definition of navigation, to allow for the possibility of navigation in organisms such as plants, which can move their body parts but are incapable of locomotion. The above definition is meant to be a broad one: it would include, for instance, the probings of bacteria, whereby they find their food.
If Albrecht-Buehler is correct, animal cells are capable of a much more sophisticated kind of navigation - visual navigation - which enables them to see objects from a long distance.
Although the navigational capacities of these animal cells are of a higher grade than those of bacteria, it would be a waste of time to distinguish the various levels of sophistication in cellular navigation without first ascertaining whether the behaviour of these animal cells is flexible - a sine qua non for possessing mental states (Conclusion F.3).
In addition to navigational capacities, animal cells also possess internal movement programs, which I describe in section 5.3 of the Appendix to chapter 2 part B. According to Albrecht-Buehler, cells' internal movement programs are not fixed: cells can over-ride them when circumstances warrant it. They are even programmed to seek information about their surroundings if they encounter more than one path they can follow. Individual cells can even co-ordinate their movements with one another (Albrecht-Buehler, 2003b).
Are the cells exhibiting flexible behaviour? To show this, we would have to show that they can acquire a new pattern of responding to a sensory stimulus (Conclusion F.3). Albrecht-Buehler's claim that the cells' internal movement programs are not fixed might seem to suggest that they are flexible. However, another more likely interpretation, hinted at by Albrecht-Buehler, is that there are different levels of control within each cell. We could think of each cell as having a master program that governs its movements. The specific functions that happen to be activated by the program may vary as environmental conditions change, but the program coding for the functions themselves, which describe the cells' patterns of responding to sensory stimuli, need not vary over time, nor does the program instructions need to change. If this is so, then the cells' behaviour would be inflexible according to the definition I am using.
In other words, even the sophisticated navigational capacities shown by these animal cells does not appear to require flexible behaviour.
Moreover, there is no evidence to date that the cells have a memory - although this is partly due to the difficulty of designing an experiment that would prove it. Nor is there any evidence that the cells have a "cognitive map" of their surroundings (Albrecht-Buehler, personal email, 30 September 2003). Since the existence of memory capacity in an organism is a necessary condition for ascribing cognitive mental states to it (Conclusion M.7), we have no warrant at present for saying that these cells have mental states.
A.8 The occurrence of navigation and guiding sensors in an organism does not provide a sufficient warrant for the ascription of mental states to it.
Case study 1: action selection in cnidaria
Nervous system
The simplest phylum of "true" animals (Eumetazoa) is the cnidaria (commonly known as coelenterates). Cnidaria have no brain, but they have a rudimentary nervous system, with neurons positioned regularly over the surface of the animal. Each neuron is in contact with its neighbours (Abramson, 1994, p. 176).
Although cnidaria do not possess a central nervous system or a brain, their nerve net permits rapid communication between cells (in some cases taking only milliseconds), over relatively long distances. In "simpler" animals, which lack neurons, communication can only occur between neighbouring cells.
Prescott (2001) considers the behaviour of cnidaria to be an important advance over that of sponges, which respond only to direct stimulation, at a very slow rate (about twenty minutes). By contrast, cnidaria exhibit "internally generated, rhythmic behavior, and co-ordinated patterns of motor response to complex sensory stimuli", allowing them to display an "integrated global response" to their environment (Prescott, 2001, pp. 5, 7). In some cnidaria, such as the hydrozoan jellyfish, the nerve net is arranged in a longitudinal circuit which supports fast attack, escape and defense reactions (Prescott, 2001, pp. 6-7).
A more complete description of the nervous system and senses of cnidaria can be found in section 5.4 of the Appendix to chapter 2 part B.
Agency in cnidaria?
Prescott considers the nervous system found in cnidaria to be a fundamental advance in the evolution of what he calls "action selection" or the problem of "resolving conflicts between competing behavioural alternatives" (2001, p. 1).
Definition - "action selection mechanism"
An action selection mechanism in an organism may be defined as a repertoire of actions, combined with the ability to select the most appropriate one for the present circumstances.
Action selection is an essential epistemic condition for the identification of mental states in an organism: without such a mechanism, the organism could not even be said to exhibit goal-directed behaviour, let alone intentional behaviour.
A.9 An organism must have an action selection mechanism before it can be said to have cognitive mental states.
Unfortunately, this requirement does not take us very far. As Prescott himself admits (2001, p. 1), action selection is part of a problem faced by all living creatures: behavioural integration, or the task of co-ordinating the activities of their parts and sub-systems. Even bacteria and plants possess this ability (Godfrey-Smith, 2001, pp. 6-7).
A.10 All cellular organisms possess an action selection mechanism of some sort.
It might be argued that any organism with the ability to select the most appropriate action for the circumstances is an agent of sorts, as it is choosing the best means of achieving its ends: "This action, not that one, will get me what I want". However, the selection need not involve the organism having its own internal means-end schema. A selection can be triggered by incoming sensory information (a stimulus). The goal achieved is what the action is for, but the organism making the selection does not need to know that, if its behavioural programs are working properly. Nor do these programs need to be flexible; the existence of an action selection mechanism is compatible with fixed patterns of behaviour. As flexibility is not required for action selection, I conclude that it can be adequately described using a goal-centred intentional stance.
A.11 The fact that an organism has an action selection mechanism does not provide a sufficient warrant for the ascription of mental states to it.
Nevertheless, Prescott (2001) contends that the cnidarian action selection mechanism represents a different kind of behaviour from that of "simpler" animals such as sponges. For Prescott, the speed and co-ordination of the way in which some cnidarians respond to stimuli represents a milestone in the history of action selection.
In some jellyfish, the nerve net is functionally divided into two relatively independent systems - one for feeding and the other for movement - which interact in neuron clusters. Others possess a single nerve net which can carry two different types of action potentials enabling either rapid escape swimming (to avoid predators), or, slow rhythmic swimming for feeding (Prescott, 2001, pp. 5-7).
Prescott likens this decentralised neural arrangement to the subsumption architecture described by Brooks (1986). A Brooksian "agent" has no central control: it is hierarchically organised from the bottom up. Control is distributed between different components, making the "agent" better able to withstand damage (i.e. more robust). Behaviour patterns are hard-wired, and sensors and actuators (which produce movement) are closely coupled, to allow rapid response times. Co-ordination between the different components is ensured by built-in timers and by having behaviour modules that can inhibit one another. Simple behaviours combine to produce more complex patterns of behaviour (Laird, 1994). According to Prescott, the functional subdivision of the nerve net into two distinct circuits for feeding and movement, which is found in some jellyfish and sea anemones, resembles the Brooksian architecture proposed for some behaviour-based robots (2001, p. 6).
I discuss Brooksian architecture at further length in section 5.5 of the Appendix to chapter 2 part B, where I argue that the specifications of Brooks' architecture make the ascription of mental states to a Brooksian "agent" redundant. A Brooksian "agent" has a very "low-tech" design. It has no internal model of the outside world, and does not engage in planning or learning of any kind. All of its behaviour is hard-wired and built-in, to ensure co-ordination and cope with unforeseen contingencies. In other words, its patterns of behaviour are fixed. If cnidaria do indeed behave like Brooksian "agents", then they cannot learn new ways of responding to unforeseen events. For reasons discussed above (Conclusion F.3), the ascription of cognitive mental states to cnidaria would then be redundant, as it would tell us nothing useful about their behaviour. A mind-neutral, goal-centred intentional stance would suffice.
Additionally, I have not been able to locate any evidence that cnidaria are capable of associative learning, which would indicate flexible behaviour. In section 5.6 of the Appendix to chapter 2 part B, I suggest follow-up experiments that could be performed in this area.
Certainly, cnidaria possess biologically significant features that other creatures lacking minds do not - in particular, a nervous system which (in some cases) allows very fast signal conduction as well as (possibly) reflexes, and enables fast attack, escape, or defense reactions. It would be a mistake to equate the behavioural repertoire of cnidaria with that of bacteria or even plants. Unlike bacteria, cnidaria are multicellular creatures, which face the task of co-ordinating their entire bodies in response to sudden changes in their environment. For instance, swimming in jellyfish requires the "synchronous, simultaneous contraction of the entire perimeter of the bell" (Prescott, 2001, p. 7). And while plants, like cnidaria, are multicellular, plants are not motile. There is no plant analogue of a rapid escape response.
However, the evidence to date suggests that the sensory capacities and behaviour modification observed in cnidaria can be adequately described using a mind-neutral intentional stance. There is no evidence that they possess an internal mechanism enabling them to modify their behaviour patterns and learn to do something new or different (Abramson, 1994, p. 38), so there is no warrant at present for ascribing mental states to them.
A.12 The fact that an organism has an action selection mechanism, sensors to guide navigation, and a nervous system with reflexes, does not provide a sufficient warrant for the ascription of mental states to it.
Case study 2: centralised action selection in flatworms
Flatworms are considered to be the most "primitive" phylum of worms. The simplest central nervous systems are found in flatworms (platyhelminthes). Flatworms also have an important evolutionary significance, as they are thought to resemble the common ancestor of all animals with bilateral symmetry (Prescott, 2001, p. 12).
Prescott considers the appearance of the platyhelminthes in the fossil record (565 to 544 million years ago) to be the next major breakthrough in the evolution of action selection, after the evolution of cnidaria. He cites research by Raup and Seilacher (1969, cited in Prescott, 2001, pp. 12 - 13) showing that trace fossils of meandering foraging trails left by the earliest flatworms, which are described in detail in section 5.7 of the Appendix to chapter 2 part B, can be generated by combining four simple behaviour mechanisms, one of which functions as a centralised conflict-preventing mechanism, of vital importance to an organism with a primitive brain and bilateral symmetry.
Should the ability of bilaterally symmetric animals to engage in centralised action selection be envisaged in mentalistic terms, as a primitive form of intelligent behaviour? Or is there an alternative, neutral description of this behaviour? Robotics researchers such as Mataric (1990, cited in Prescott, 2001, p. 13) have pointed out that the first three mechanisms imputed to the earliest flatworms are identical to those used by a robotic wall-follower, while the fourth can be generated simply by transferring control of staying close to the wall from one sensor arm to the other.
It has been argued above (Conclusion L.4) that there can be no "true" learning (and hence no mental states) in an organism unless its behavioural response patterns are flexible. The fact that flatworms' action selection mechanisms (unlike those of cnidaria) are centralised does not imply that they are flexible.
A.13 The occurrence of centralised action selection, sensors and a central nervous system in an organism does not provide a sufficient warrant for the ascription of mental states to it.
Why fine-tuning is required for agency
We have already seen that many animals are capable of learning to perform certain actions for the sake of the consequences they learn to associate with them (e.g. the attainment of a reinforcement). Most psychologists refer to such behaviour as operant behaviour, but Abramson calls this kind of behaviour "instrumental conditioning", and reserves the term "operant behaviour" for a special sub-set, which he defines as "the ability to operate some device - and know how to use it, that is, make an arbitrary response to obtain reinforcement" (1994, p. 151, italics mine). As we saw above, some scientists find the distinction between instrumental and operant conditioning unacceptably vague. Recently, however, Abramson provided me with more specific examples of behaviour conforming to his more restrictive definition:
For example, we know that rats can be taught to press a lever in various directions and with various degrees of force. They can also be trained to run down an alley with speeds selected by the experimenter... (personal email, 2 February 2003, italics mine).
I would suggest that Abramson's description of operant behaviour can be re-expressed using the concept of fine-tuning. What the rats in the above examples are doing is confining some parameter that describes their movement (speed or force exerted) within a narrow range.
It would be a misunderstanding to infer that undergoing operant conditioning involves learning to move in a new way. There is no need for such a high degree of creativity on the animal's part; all it to needs to do is select an existing motor pattern from its repertoire and stabilise or confine it within a narrow range.
Finally, it needs to be stressed that fine-tuning behaviour is flexible and internally generated, because it arises through a learnt association between the performance an action and the obtaining of a goal (e.g. a reward, or the avoidance of punishment) (see Conclusion L.8).
Definition (first draft) - "operant conditioning"
Definition - "fine tuning"
Fine tuning refers to an individual's act of stabilising a basic motor pattern at a particular value or confining it within a narrow range of values, in order to achieve a goal that the individual has learned to associate with the action. Fine-tuning can be regarded as a (learned) refinement of action selection.
Operant conditioning describes a process of instrumental conditioning in which an animal learns to fine-tune its movements in order to obtain some goal.
The foregoing definition of operant conditioning is only a preliminary one, whose main purpose is to provide researchers with a relatively straightforward way of distinguishing operant learning from other forms of instrumental conditioning. I shall refine the concept later in this chapter.
The notion of "fine-tuning" can be tied to another related concept: that of control. If an animal can adjust and fine-tune its responses to a variable stimulus, then we can say that it has control over its responses, and is hence able to engage in operant behaviour.
An animal that cannot fine-tune its responses to a stimulus, lacks control over them and is only capable of instrumental behaviour at best.
Earlier in this chapter, I argued that mental states such as beliefs and desires are appropriately identified through the performance of intentional acts. In ordinary discourse, the notion of acting intentionally is closely tied to the notions of trying and control. I would like to make the stronger claim, that the notion of agency presupposes that of control, which is manifested in fine-tuning behaviour. (It will be argued in Part C of this chapter that there are at least four distinct kinds of behavioural manifestations of "fine-tuning".) Specifically, I assert that an organism must be capable of fine-tuning its bodily movements before it can be identified as an agent.
To see why, it will be helpful to examine Carruthers' (2004) argument against the possibility of attributing minds to animals, solely on the basis of what they have learned through conditioning:
...engaging in a suite of innately coded action patterns isn't enough to count as having a mind, even if the detailed performance of those patterns is guided by perceptual information. And nor, surely, is the situation any different if the action patterns aren't innate ones, but are, rather, acquired habits, learned through some form of conditioning.An organism with an action selection mechanism has a fixed repertoire of actions, from which it is able to select the most appropriate one for the present circumstances. When an organism undergoes classical conditioning, its action selection mechanism learns to do something novel: the presence of a neutral stimulus (e.g. the sound of a bell) causes it to automatically select a certain action (e.g. a dog learns to salivate when it hears a bell), which it now associates with one of its built-in goals (e.g. food, which naturally makes it salivate). To explain a given occurrence of this behaviour, all we need to specify is some sensory information (about the presence of the neutral stimulus) and a built-in goal (food). We have no reason to suppose that the link the organism makes between the two events (bell -> food) takes the form of a belief.
In instrumental conditioning, the organism learns that performing one of its repertoire of actions in particular circumstances will enable it to achieve one of its goals. Here, it might seem natural to suppose that the organism performs the action because it believes that by doing so, it can get something it wants. But there is a simpler explanation: its past experiences (encoded as information in its brain) bias its internal action selection mechanism, causing it to perform the action that will in fact obtain for it something it wants (its goal).
Thus although the behaviour of the animal in each of these cases is flexible (insofar as its internal "behavioural program" is modified), a goal-centred intentional stance appears adequate to explain the animal's behaviour.
The upshot of these arguments is that in the absence of fine-tuning behaviour, possession of an action selection mechanism is insufficient to generate behaviour that clearly manifests intentional agency, even if the behaviour is flexible (e.g. learned behaviour). We also saw earlier that behaviours such as directed movement and navigation do not require fine-tuning, as they can be generated by fixed patterns of behaviour:
A.14 An organism must be capable of fine-tuning its bodily movements before it can be identified as having cognitive mental states.
L.15 Neither an animal's capacity to undergo classical conditioning nor its ability to learn from instrumental conditioning, per se, warrant the ascription of cognitive mental states to it.
The question of whether fine-tuning is a sufficient condition for the presence of cognitive mental states (such as beliefs) will be discussed below.
Which organisms are capable of fine-tuning?
Case study 1: The nervous system of cnidaria

Cnidaria (commonly known as coelenterates) are the most "primitive" phylum of "true" animals (Eumetazoa). They have a rudimentary nervous system, described by Abramson:
Neurons are located regularly over the surface of the animal... The propagation of a nerve impulse is not transmitted along a linear chain of neurons, but radiates from its point of origin... The effect of such an arrangement is that a stimulus applied to any part of the animal will be directed to all parts, much like sticking your finger in a cup of jello will make the whole mass move... Such a system is not conducive to fine control of motor movements (1994, p. 176, italics mine).
If we wish to identify fine-tuning, we must look for it in other phyla of "true" animals - i.e. those with central nervous systems.
A.15 Only organisms with central nervous systems are capable of fine-tuning their bodily movements for the performance of intentional acts.
Combining this with conclusion A.14, we obtain the result that we can only ascribe cognitive mental states to organisms with central nervous systems. This is identical with the conclusion we reached in the previous section (conclusion B.6).
Case study 2: Operant conditioning in C. elegans worms?

Earlier, I mentioned two recent studies (discussed in section 4.3 (case study 3(d)) of the Appendix to chapter 2 part B) suggesting that C. elegans worms may indeed be capable of modifying their responses to obtain a reinforcement. Although the studies produced some conflicting results, they indicate how one might proceed in attempting to verify operant behaviour - as defined by Abramson - in a species of animal.
First, it is important to construct a complete pictorial catalogue of the behavioural patterns (in particular, movement patterns) of that species - i.e. an ethogram.
Second, it is essential to learn what attracts them and what repels them.
Third, the animals have to be able to control (or fine-tune) at least one of their patterns of behaviour, in order to obtain a "reward" or avoid a "punishment".
Some questions still need to be addressed. Exactly how are we supposed to identify fine-tuning? How do we know if an organism is exercising "control" over its patterns of behaviour? I will discuss these matters further in the next section, in connection with Dretske's account of belief.
Finally, we come to the question of whether the occurrence of "fine-tuning" or "control" in animals indicates intentional agency on their part. While the word "control" easily lends itself to a mentalistic interpretation (i.e. an agent-centred intentional stance), it is by no means clear why the behaviour of an animal that can learn how to stabilise its motor patterns in order to obtain a goal, requires an agent-centred description. At the very least, the notion of "fine-tuning" needs to be developed at further length before it can support any attribution of beliefs to animals. In the following sections, I examine two key concepts - the concept of representation and the concept of self-correction - which can help us to make the notion of "fine-tuning" more precise.
It has been argued that beliefs are essentially intentional: to have a belief is to have a belief about something. Also, because a belief is a mental state, a belief-holder must possess a mental representation of the intentional object of her belief. But what does it mean to represent something?
Case study - can bacteria represent magnetic north?

Schwitzgebel (2003) cites a case (taken from Dretske, 1988) which may suggest that certain bacteria are capable of forming representations of their environment, if we view representational systems as systems that track features of their environment. For philosophers who adopt a representationalist account of belief, this raises the question of whether bacteria are capable of having beliefs. The bacteria in question have compartments called magnetosomes, which contain particles of magnetite.
Organisms, especially mobile ones, generally need to keep track of features of their environment to be evolutionarily successful. Consequently, they generally possess internal systems whose function it is to covary in certain ways with the environment. For example, certain marine bacteria contain internal magnets that align with the Earth's magnetic field. In the northern hemisphere, these bacteria, guided by the magnets, propel themselves toward magnetic north. Since in the northern hemisphere magnetic north tends downward, they are thus carried toward deeper water and sediment, and away from toxic, oxygen-rich surface water. We might thus say that the magnetic system of these bacteria is a representational system that functions to indicate the direction of benign or oxygen-poor environments... In general, an organism can be said to represent P just in case that organism contains a subsystem whose function it is to enter state A only if P is true, and that subsystem is in state A (Schwitzgebel, 2003).
But are the bacteria representing anything here? Procyshyn (2001) argues that they are not. A representational account has to do some extra explanatory work that a purely causal account cannot; otherwise it is redundant. An example from Procyshyn illustrates why: when someone taps my bent knee with a mallet, a neural pathway fires, causing my leg to jerk and straighten - a reflex process over which I have no control. Now, we could say that the external stimulus (mallet) is represented by my neural pathway, and that the function of this representation of to straighten my leg. Or we could explain the reflex in purely causal terms. Given that both accounts describe what goes on equally well, we should prefer the simpler, causal account.
Following Dretske, Procyshyn then proposes that "a representation's function is to supplement a causal chain of events" (2001, p. 134, italics mine) and that "the function of a representation is to encode, use or deliver information about the external world that is pertinent to the representing organism's survival" (2001, p. 134). Since, for these bacteria, the downward direction of the benign environment correlates with the direction of magnetic north detected by the bacteria's sensors, we can say that an external state of affairs is correlated with an internal state of the bacteria, whose biological function is to use, encode or deliver information to the bacteria that assists in their survival. Thus Dretske considers that the bacteria have a genuine representational capacity (although, like most representationalists, he does not credit them with beliefs).
But as Dretske points out, if we hold a bar magnet over these bacteria, they align themselves with the magnet, not with magnetic north. What the bacteria are doing is aligning themselves with the nearest magnet. Procyshyn comments:
...[G]iven the manner in which the magnetosome represents, it is incapable of misrepresenting. Since the magnetosome appears to represent magnetic north proximally, there is no way for the magnetotactic sensor to misrepresent. No malfunction is possible here (2001, p. 135).
Procyshyn argues that a representation, by definition, is something that can be right or wrong, true or false. If the bacteria are not capable of mis-representing, then we should not speak of them as representing either. And if they cannot be said to represent, then the question of bacterial beliefs does not arise. Since a capacity to represent is a pre-requisite for having a belief, we can formulate a minimal criterion for the ascription of beliefs to organisms:
R.1 A necessary condition for the ascription of beliefs to an organism is that it be capable of mis-representing events occurring in its surroundings.
Bacteria, like all other organisms (except viruses) are certainly capable of encoding a variety of information about events in their environment. However, we need to be careful not to describe their capacities in philosophically inappropriate ways, thereby generating pseudo-problems.
Dretske: why operant conditioning is evidence for belief and agency in animals
Earlier, when discussing Dretske's (1999) article on machines, plants, animals and agency, I suggested that there were actually two criteria in his article which he used to distinguish between organisms with and without beliefs. I examined his first criterion of learned flexible behaviour, and found that it could not account for three cases (conditioning of the autonomic nervous system (ANS) in an astronaut, of leg withdrawal in headless cockroaches, and of flexion of the hind legs within the spinal cords of paralysed rats) where learned flexible behaviour appears to occur in the absence of beliefs. Additionally, Dretske seemed to employ the terms "mindless", "unlearned" and "hard-wired" inter-changeably.
However, Dretske used another, more rigorous criterion for distinguishing organisms that have beliefs and exhibit agency from those that do not: the capacity to undergo operant conditioning, a form of learning which he links to the notions of representation and control:
Dretske Mark II
Learning of this sort (operant conditioning) consists in harnessing these internal representations to control circuits so that behaviour will occur in the external conditions on which its success depends
(1999, p. 10, italics mine).
How are we to understand Dretske's claim that operant conditioning can be understood in terms of internal representations linked to a controlled behavioural response? Two readings are possible. On a "maximalist" reading (which I shall defend below in my model of operant agency), an organism with beliefs is one that can use its internal representations of its environment, acquired through learning, to control its surroundings. The verb "control" is here meant to describe an action, performed by an agent. (On this account, the concept of agency needs to be further developed.) An alternative "minimalist" reading (which appears to be Dretske's own view) is that operant agency is simply behaviour which is controlled by an organism's internal representations of its environment. Here, "controlled by" simply means "caused by".
Dretske's account of representation is a thorough-going naturalistic, causal account, which eschews appeal to "interpreters" as the arbiters of what counts as a representation. Representations, on his account, are indicators which carry information about lawlike connections (say, between As and Bs), but they are something more. Representations, unlike indicators, can be mistaken, because they have a function which they can fail to perform. More precisely, representations are indicators whose natural function is to indicate as they do, because doing so confers a selective advantage on the organism possessing them. Representations, unlike other natural indicators, are not hard-wired: they acquire a function or meaning for an animal only when the animal learns what they indicate. An animal's learning history imparts a meaning to its experiences. Belief-type representations are recruited as causes of bodily movements in an animal because the animal learns what they indicate. Thus beliefs are both reasons and causes of actions (Ryder and Martin, 1999, pp. 5-7; MacFarlane, 2003; Pitt, 2002). This explains Dretske's (1999) contention that while behaviour may have a meaning, purposeful acts are governed by their meaning, insofar as events cause an animal to behave in a certain way, by virtue of their meaning.
Thus in Dretske's case of the bird that shuns the Viceroy after tasting a similar-looking noxious Monarch butterfly, the bird's internal representation causes the avoidance behaviour precisely because it means something about its external environment (i.e. that a certain kind of butterfly is present - the sort of butterfly the bird, after its unpleasant experience, wants to avoid).
It would be beyond the scope of this thesis to adjudicate between competing theories of mental representation (summarised in Pitt, 2002). What is important here is that in Dretske's account (unlike Abramson's, which was discussed above) internal representations play a causal role in operant behaviour. It is also significant that Dretske's account highlights the evolutionary history of representations, which confer a selective advantage on their possessor.
Dretske's account of belief: a critical evaluation

Despite these positive features, I believe that Dretske's account of belief is not powerful enough, on its own, to distinguish organisms with beliefs from those without.
First, his account fails to distinguish operant behaviour from mere instrumental conditioning, which (Conclusion L.14) can occur even in the absence of belief. We looked at three such cases above. Dretske might claim that in these cases, the systems involved are not forming representations. But in fact, the astronaut's ANS is representing its external environment, and its function is undeniably a natural, biological one. Moreover, some kind of learning is going on: in space, the astronaut's ANS learns not to compensate for loss of blood to the brain when she stands up, and once the astronaut returns to earth, her ANS has to re-learn this skill. Why does this not qualify as operant conditioning?
The conclusions we reached in section 5 provide us with a ready answer to this question: because the astronaut's ANS is not engaging in fine-tuning behaviour, it must be undergoing instrumental rather than operant conditioning. However, the notion of "fine-tuning" does not figure in Dretske's (1999) account of belief.
To add to the confusion, Dretske's own example of the bird that shuns the Viceroy is not a proper example of operant conditioning, strictly speaking. According to Dretske, the bird believes that the Viceroy tastes unpleasant, and I would certainly agree with him that this ascription of belief to the bird is correct and natural. But in this case, the bird can hardly be said to be engaging in operant behaviour, properly speaking: it is merely avoiding an object that resembles a noxious stimulus. (Abramson would classify this as instrumental conditioning.) One could explain the bird's change of behaviour using a mind-neutral goal-centred intentional stance, according to which memory can be understood simply as stored information, and learning as a process by which the bird, as an educable animal, acquires new information that enables it to alter its behaviour patterns.
Finally, Dretske's minimalist account fails to explain why behaviour controlled by representations has to be envisaged in mentalistic terms. For Dretske, "controlled behaviour" appears to be synonymous with behaviour caused by representations - defined as learned connections between As and Bs, which serve a biological function. But causation per se is not a mentalistic notion. Likewise, Dretske's notion of representation is too weak to sustain mentalistic inferences. For him, the decisive features of representations are that they carry information which indicates something about the world, they confer a selective advantage on their possessor and they are learned. As we have seen (Conclusions I.1, B.5, I.3, L.11, L.13, L.14), these properties are necessary conditions but not sufficient warrants for the ascription of mental states.
R.2 The presence in an organism of Dretskean representations, defined as indicators acquired through learning which serve a biological function, does not provide a sufficient warrant for our being able to ascribe cognitive mental states to it.
Despite these problems, I believe Dretske's account of belief lies very close to the truth, because it brings together vital ingredients of belief: flexible behaviour, learning, internal representation, and the possibility of error (mis-representation).
Dretske could have made his account more robust by including a discussion of fine-tuning, which as we saw is an essential condition for the ascription of beliefs to an animal (conclusion A.14). Dretske's representational account of belief complements the account of fine-tuning we developed in section 5. We have seen that behaviour controlled by representations does not have to be envisaged in mentalistic terms, but one might argue that fine-tuning behaviour controlled by internal representations is indeed the kind of behaviour that could serve to manifest underlying beliefs. It seems natural to say that an animal whose internal representations cause it to stabilise one of its motor patterns within a specified range, in order to attain a goal, is doing so as a means to an end: it believes that by fine-tuning its movements, it will get what it wants.
I would suggest that Dretske's account could also be strengthened by adopting a richer notion of representation. In my model of operant agency in insects in part C, I argue that representations have a distinctive internal structure. In describing this structure, I draw upon several themes that have been discussed in this section: action selection, fine-tuning, control, trying, associations, means and ends (goals). I also argue that this rich structure also allows certain kinds of representations to serve as beliefs through which agents control their actions.
It remains to examine one more criterion that has been proposed as the hallmark of belief: the possibility of self-correction.
Beisecker on belief
Whereas for Dretske (1999) the salient feature of beliefs is that they are internal representations, acquired through learning, which bring about changes in organisms' behaviour, for Beisecker (1999) the defining quality of beliefs is that they can be correct or mistaken. As he puts it:
...the hallmark of intentional states is their susceptibility to evaluation. For instance, a doxastic (or belief-like) state can be correct or mistaken depending upon whether or not some state of affairs, identified as its content, actually obtains. Believers are beholden to the way things are. Similarly, conative states (goals, desires, and other "pro-attitudes") may be fulfilled or unfulfilled, depending upon the satisfaction of some content condition. So intentional states are those that are associated with conditions of satisfaction or fulfillment (1999, p. 283).
The notion that susceptibility to evaluation is a distinguishing feature of intentional states may seem strange to people who regard mental states as quintessentially incorrigible - an idea expressed in the naive claim that I cannot be wrong about my own mental states, whatever else I am wrong about. Even if this claim were correct, it would in no way weaken Beisecker's point, which is that however well-acquainted I may be with my mental states, they are about something. It is in terms of their aboutness that they are vulnerable to evaluation as right or wrong, satisfied or unsatisfied.
The same goes for the Aristotelian claim that the senses cannot be deceived about their special objects (e.g. for sight, colour). We do not simply see colours; we see coloured things. And it is when evaluating the intentional object of our vision that we are capable of erring - a point well-understood by Aristotle (De Anima 2.6).
For Beisecker, animals that are capable of a special kind of associative learning - operant conditioning - qualify as belief-holders. According to Beisecker, animals capable of operant conditioning should be regarded as having beliefs because not only are they educable, but they can make errors which they subsequently try to rectify.
Beisecker argues that animals capable of operant conditioning possess a special non-biological kind of intentionality. Unlike other organisms, whose ends are completely biological, these animals can be said to have expectations that their responses to certain kinds of events will bring about certain outcomes. Insofar as a creature engages in behaviour expected to bring about a certain outcome, we may regard that outcome as one of its goals. Beisecker call these goals non-biological, because we can identify them without having to know anything about the creature's biological evolution, which has determined its built-in ends through natural selection.
Of course, expectations may be disappointed: a creature may make errors of commission (when an animal's expectation - say, of obtaining food - is activated and it responds, but the expected consequence does not eventuate) and errors of omission (when the animal fails to respond because its expectation is not activated, but in fact, the response would bring about a desired consequence). But because creatures continually revise their expectations, they can be said to possess a kind of critical rationality:
Insofar as they are disposed to revise their expectations in the wake of the errors described above, educable creatures would be disposed to take steps to avoid similar mistakes in the future. There is of course no guarantee that these revisions will yield future success. The point is just that creatures displaying this sort of educable capacity would take expectation correctness or aptness to be a regulative ideal, at least in the sense that they are disposed to revise error-prone expectations while leaving correct expectations as they are... Since they can be evaluated as having gotten things right or wrong, we are justified in crediting these creatures with some sort of intentional capacity (Beisecker, 1999, p. 303).
In other words, animals that can undergo operant conditioning can be said to be capable of getting things right, because they can (and often do) get them wrong, and they revise their expectations and their responsive behaviour when they are wrong. On Beisecker's account, the capacity to self-correct one's mistakes is a sine qua non for having beliefs. I have argued above that we can adopt a mind-neutral goal-centred intentional stance, when explaining the behaviour of that an organism lacking beliefs. A minimal mind must therefore be capable of entertaining at least some beliefs, about which it can be right or wrong. We can propose the following normativity criterion:
N.1 An organism must be capable of self-correcting behaviour before it can be said to have cognitive mental states.
Beisecker's proposal deftly handles counter-examples associated with organisms that behave maladaptively, such as the bacteria containing magnetsosomes, which were discussed above. These bacteria should not be described as being "in error" when they move toward the bar magnet instead of the bottom of the water, because they cannot "tailor their responsive dispositions to their particular surroundings" (1999, p. 298), and hence do not qualify as holders of expectations or beliefs. Bacteria do not try to rectify their maladaptive responses, and it was found that a causal account could account for their behaviour.
Beisecker's model of animal behaviour makes a novel prediction that a purely causal, non-mentalistic account does not: animals that are capable of operant conditioning will revise their mistaken expectations and try to correct their mistakes. However, this "prediction" cannot be considered scientifically productive unless it can be fleshed out in behavioural terms. What kind of behaviour counts as self-correction, and why? Only if we can answer this question will we be able to identify animals with beliefs and desires. In part C, I propose a detailed answer, in the context of operant agency.
At this point, one might ask whether motor fine-tuning could count as self-correcting behaviour. However, there are two fundamental problems with this proposal.
First, fine-tuning has to do with confining a motor pattern within a specified range. By itself, it cannot tell us how to rectify any deviation outside the desired range.
Second, fine-tuning deals with movement, whereas self-correction pertains to revision of one's beliefs, which means adjusting one's internal representations. In other words, self-correction cannot be cashed out in purely external terms; it is an internal process as well.
As we have seen, there are differing schools of thought as to whether operant conditioning should be envisaged in mentalistic terms or purely causal terms. One phenomenon - blocking - has been proposed by Beisecker as an example of behaviour that expectation-generating animals would engage in. Blocking is a highly unusual prediction of one model of associative learning - the Rescorla-Wagner model - whose central ideas are usually expounded using mentalistic terminology.
The basic principle of the Rescorla-Wagner model is that "the amount of conditioning depends on how surprising the association between the CS and US is. The more unexpected or surprising the US, the more conditioning will occur" (Lipp, 1998, italics mine). While the model is not able to explain all phenomena connected with classical conditioning, it is still regarded as "the 'best' theory of classical conditioning" (Jackson, 2002).

The model readily explains the phenomenon of blocking, in which animals that have been trained to associate a conditioned stimulus (e.g. a bell tone) with an unconditioned stimulus (e.g. an electric shock) will fail to associate a second conditioned stimulus (e.g. a red light) with the unconditioned stimulus, if the latter is subsequently presented along with the original conditioned stimulus (the bell tone). The model explains this curious fact as follows: initially, the animals associate the bell tone with the electric shock because the co-occurrence of the two events surprises them. The addition of a second conditioned stimulus generates no new unpleasant surprises for the animals, so they do not make any new associations. What the model seems to suggest is that animals undergoing operant conditioning form new expectations of what will happen, and hence have beliefs. If this is correct, the phenomenon of blocking could be used to distinguish those animals which are capable of forming expectations from those which are not (Beisecker, 1999, pp. 298-299).
There are two good reasons for caution here, which I discuss in greater detail in section 7.1 of the Appendix to chapter 2 part B. First, most kinds of animals that are capable of operant conditioning have not yet been shown to exhibit blocking: there have been no confirmed reports of blocking in worms, and its occurrence even in so-called "higher" invertebrates (honeybees) remains controversial.
Second, blocking may turn out to be explicable in non-mentalistic terms, in any case. Some scientists use the term "expectation" in a mind-neutral sense, to denote a memory retrieval function that results from former learning. Some even attempt to explain blocking as the result of peripheral sensory integration (Menzel, personal email communication, 21 July 2003). We have already seen that sensory capacities and memory can be explained by adopting a goal-centred intentional stance. If many scientists believe that these capacities are sufficient to explain blocking, then (pace Beisecker) it would be unwise to invoke blocking as evidence of mental states.
L.16 The occurrence of blocking in an organism does not provide a sufficient warrant for our ascription of cognitive mental states to it.
I also present evidence in section 7.2 of the Appendix to chapter 2 part B, for so-called "higher-order" forms of associative learning (not only blocking, but also overshadowing, sensory pre-conditioning (SPC) and second-order conditioning (SOC)) in fruit flies, before reaching the following negative conclusion:
L.17 The occurrence of higher-order forms of associative learning in an organism do not, taken by themselves, warrant the conclusion that it has cognitive mental states.
Table 2.4 - Which organisms meet the requirements for intentional agency?
| Category of Properties | Required Property for Agency | Bacteria | Protoctista (e.g. amoebae, algae, seaweeds & paramecia) |
Plants | Cnidaria (coelenterates) |
Worms | Insects |
| Biological | Central nervous system (to allow fine-tuning) |
No | No | No | No, but nerve net permits rapid signal conduction | Yes | Yes |
| Sensory | Sensory capacities | Yes, but locomotion-driven | Yes (slow, chemical-based) |
Yes (slow, chemical-based) |
Yes | Yes | Yes |
| Memory | Procedural memory | No (chemical only) | No (habituation) |
No (habituation) |
No (habituation) |
Yes | Yes |
| Flexible behaviour | Internally generated flexibility | No. Flexible behaviour is externally driven. | No. Flexible behaviour is externally driven. | Probably not, but very complex movements. |
Not known, but possesses fast escape mechanisms. |
Yes. | Yes. |
| True learning | Operant conditioning | No | No (habituation only) |
No (habituation only) |
No (habituation only) |
Not known. Capable of classical and instrumental conditioning |
Yes |
| Controlled action | Motor fine-tuning | No | No | No | No | Not yet known | Yes (see Part C) |
| Representation | Indicators w. biological function; acquired by learning; capable of making mistakes | No | No | No | Not known, but some jellyfish can see |
Yes | Yes |
| Normativity | Self-correcting behaviour | No | No | No | No | Not known | Yes (see Part C) |
Some of our key terms still lack a complete definition (operant conditioning) or adequate identifying criteria (self-correcting behaviour). I shall attempt to remedy these problems in part C. In the meantime, insects (at least, some of them) appear to satisfy our requirements intentional agency, worms might do so, while cnidaria and plants probably do not.