
Some ecological aspects of Chysomyxa
abietis (Unger)
epidemiology
Antti Uotila 1, Tianfu Wang 2, Limei Wang3
Key Words: Picea abies, needle rust, Chrysomyxa abietis, needle cast, chlorosis, spore trap, hyperparasite, E. caricis.
Authors´address: 1 Hyytiälä Forestry
Field Station, Helsinki University, Hyytiäläntie 124, 35500 Korkeakoski,
Finland. Email: auotila@hyytiala.helsinki.fi;
2
Department
of Plant Biology, Helsinki University, PL 28, Koetilantie 5, Helsinki,
Finland. Email: tianfuw@evitech.fi;
3
The program of International Forestry, Tampere Polytechnic, Rasinkatu 10D
107, 01360 Vantaa, Finland. Email: limeiw@hotmail.com.
Spruce needle rust Chrysomyxa abietis (Wallr.) Unger causes temporal epidemics on spruce needles. Picea abies ( L.) Karst, Picea sitchensis (Bong.) Carrière, Picea rubens Sarg., Picea pungens Engelm. and Picea engelmannii Parry ex Engelm. can be the hosts of C. abietis (Murray, 1953).
C. abietis produces only telia and basidiospores ripening on underside of second-year spruce needles. No alternative host is required in the fungal life cycle (Peterson 1963; Grill et al., 1978). Golden yellow, pustule-like teleutosori develop on last-year-growth needles. The diseased needles remain on the shoots over winter and later cast after basidiospores are produced and released.
In general pustules of the fungus are orange, up to 10 mm long. The pustules consist of chains of orange or red-brown teliospores (14-42 x 9-16 m m) (Anonymous). The mycelia of the fungus are hyaline with orange or yellow droplets (Grill et al., 1984).
Basidiospores infect the young developing needles and hyphae penetrate into the needles through stomata (Grill et al., 1980). The infected needles turn yellow in some weeks after infection. The diseased needles die and fall after basidiopsores release. Current-year needles remain on shoots (Grill et al., 1978). In Norway, crown density of Norway spruce stands in 1988-1996 has been decreased in average 0.45% annually and mainly caused by C. abietis (Solberg, 1996). The rust can be closely associated with spruce needle cast fungus Lophodermium piceae (Fuckel) Höhn (Lehtijärvi, 1998).
In late summer telia start to appear on the undersides of the needles, but they do not complete their development until next spring. Teliospores germinate without dormancy and produce promycelia where basidiospores are formed (Cummins and Hiratsuka 1991). Basidiospores are dispersed by means of rainfall, water splash or wind. Low temperature and radiation; high relative humidity and rainfall enhance the liberation of basidiospores (Collins, 1976).
Eudarluca caricis (Fr.) O. Eriksson (anamorph Darluca filum (Biv.-Bern. Ex Fr.) Cast) has been found parasiting spruce rust pustules (Kujala, 1950; Grill et al., 1981). Mycoparasites could decrease the spore production.
The aim of this study was to collect the basic data of current spruce
needle rust epidemics. This gives valuable information for future research
planning connected to needle rust.
2. Materials and methods
The study was carried out in the summers 1998 to 2000 at Hyytiälä Forestry Field Station of Helsinki University, which is located at Juupajoki municipality in central Finland (24° 17¢ N, 61° 50¢ ).
A small scale inventory of disease were made to get numerical data of disease incidence in 1998. Two 500-meter inventory lines were marked along two intersecting transacts near Hyytiälä Forestry Field Station, where mixed-age Norway spruce stands were growing. Five sample plots (radius = 5,64 m, 100 m2), 50 m apart, in both line were measured. Spruce tree diameter at breast height and the occurrence of C. abietis was assessed. The spruce seedlings (< 4 cm bhd) were assessed diseased or healthy. The criteria for fungal infection was to detect any yellow pustule of C. abietis on Norway spruce needles. Five random sample trees were selected for assessing the infection rate of last-year needles. The five sample trees were chosen by standard of the mean diameter of the total measured trees. The branches growing on the middle of the stem were cut and last-year-growth shoots were then removed to count the number of healthy and infected needles.
The microscope preparates from telia were made with special way. By using stereomicroscope the pustule was cut carefully along surface of the pustule, and transferred to water drop on glass slide. The slide was not covered for avoiding to destroy the structure of basidiospores.
In order to assess liberation of basidiospores, a 7-day recording volumetric spore trap (10 L air/min.) was set up under the diseased spruce branches in a 25 years old Scots pine stand, where young Norway spruce was growing under the canopy. The spore trap was mounted in the vicinity of forest meteorological station. The initial starting time of spore trapping was at 14:00 p.m. on 10th June, 1998. Melinex hyaline tape 200 gauge was marked for each 3 hours and fixed on the disk. Examination of the rust spores under a microscope was carried out every 7th day. The Melinex tape was slightly unfixed from the disk and placed on a clean paper and the surface of trapped spores must be upwards.
The tape was divided in an interval of 6 mm which was represented spore collection of three hours. The piece of cut tape was placed on the counting area of Bürker chamber (0.0025mm2 /column), a cover glass was then placed on the tape with one drop of distilled water. By using light microscope, the number of yellow basidiospores where then counted for representing liberation of basidiospores at each 3 hours.
The measuring sensors for environmental data were below 10 m distance from the spore trap. Temperature, rainfall, radiation and wind speed were recorded in five-minute intervals continiously. The values of relative humidity at 3 m height were calculated based on ventilated dry and wet temperatures.
3. Results3.1. Characteristics of Chrysomyxa abietis
Telia were hypodermal, orange or reddish brown, growing in pustules of diseased spruce needles. Pustule length varied from 1.8 mm to 7 mm (table 1). Young pustules were covered by needle epidermis and later the needle epidermis were broken and orange pustules exposed on underside of the diseased needles.
Teliospores, oblong or oval with truncate base, aseptate, hyaline with thin walls in short chains which the upper ones were wider than the down ones with size of 30-7.5 x 20-7.5 m m (mean 17.5 x 11 m m). Teliospores were full of yellow droplets.
On the top of teliospores one or several light protrusions occurred and soon extended to promycelia (fig. 1). Mature mycelia were hyaline containing yellow droplets, growing in mesophyll of spruce needles.
Promycelia produced yellow basidiospores, 5-12.5 x 7.5-17.5 m m (mean 10.1 x 12.6 m m) (fig. 2) Basidiospores germinated when they were placed on the agar media. Initial hyphae were growing from tips of the spores. On the malt agar the fungus produced secondary basidiospores, but later the mycelia died (fig. 2). After basidiospores having been released, the pustules of the rust became dark-red or black, the needles, finally casted from shoots. In case of severe infection, almost all needles were lost from the last-year-growth shoots, only new-growth shoots and needles were remained. Next year in these branches, Sirococcus conigenus (DC.) P. Cannon and Minter can be found killing the shoots finally.
 |
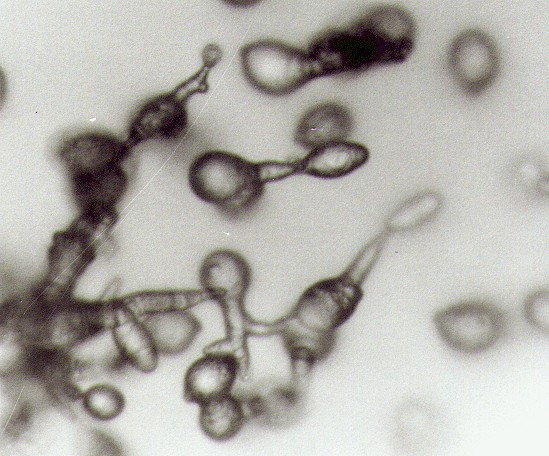 |
| Fig. 1. Promycelia are growing on teliospores. Scale bar = 7 m m | Fig. 2. Basidiopores germinate and secondary basidiospores are growing in malt agar. Scale bar = 15 m m |
Table 1. Size of teliospore, basidiospore and pustule of the rust
|
|
|
|
|||||
|
|
|
|
|
|
|||
|
|
|
|
|
|
|||
|
Min.Max. |
0,3-6,2 |
7,5-30 |
7,5-20 |
10-15 |
7,5-15 |
||
3.2. Infecting rate of Norway spruce
The result of inventory for fungal infection of Norway spruce trees and seedlings under the canopy in 1998 and 1999 was different. In 1998, 63% of spruce trees and 86% of spruce seedlings were infected. In 1999 no severe infections occurred, 10% of spruce trees and 51% of spruce seedlings were infected (table 2). On June 1998 it was rainy and relative humidity was 364 hours over 90 % in June. In June 1999 relative humidity was over 90 % only 168 hours. The humidity was enough to spore germination only on night time then and it seemed not to be enough for successful infections (fig. 3). Chrysomyxa abietis also infected big trees, e. g. a 250-year-old spruce tree was slightly infected in the garden of Forestry School at Kuru. The disease normally occurred in the lower part of crown on big trees, however it was found over 20 m high in moisture microclimate conditions. The severely infected trees cast their needles in the middle of July.
Table 2. The rate of natural infections of spruces
and seedlings at Hyytiälä in 1998 and 1999
|
trees |
seedlings |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
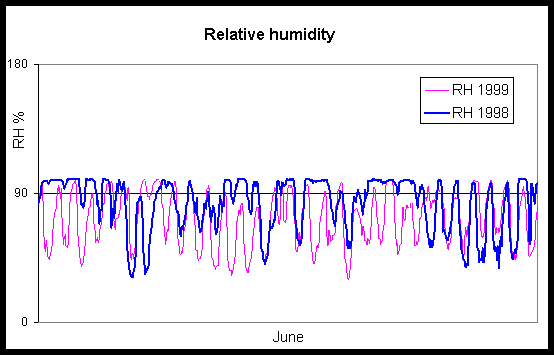
Fig. 3. Relative humidity in June of 1998 and 1999 at Hyytiälä forest meterological station
3.3. Infections in 1998-year needlesThe rate of infected needles on the sample trees varied between 7.4 to 41.1 % (table 3). Most diseased needles cast after the fungus having released basidiospores in the middle of July . Only new-growth needles remain on top of shoots.
Table 3. Rates of infections in 1998-year needles
|
trees |
examined shoots |
needles |
infected needles |
infected needles |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
3.4. Parasitic fungus of C.abietisBlack pycnidia of Eudarluca caricis (Fr.) O. Eriksson occurred in rows in the rust sori (fig.4). Pycnidia were round, up to 0,16 mm in diameter. On the top of pycnidia, irregular openings occurred for releasing conidia. Conidia were hyaline with small gelatinous caps, fusiform, one septate where two sides of the spore swapped in. Individual measurement of the spores was 5- 10 x 10-20 µm (mean 6.1 x 16.5 µm) (fig. 5). Conidia of the fungus germinated and hyphae grew in malt agar initially, but the mycelia died after the germinating conidia were transferred to the new malt agar media.
 |
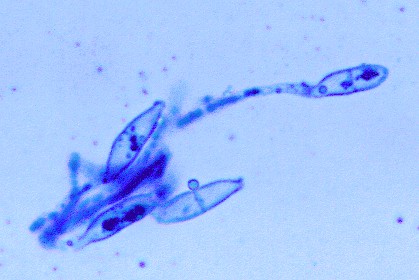 |
| Fig. 4. Black pycnidia of E. caricis growing on the surface of rust sori. The upper rust sori are free of infection | Fig. 5. Conidia of Eudarluca caricis, stained by Cotton blue. Scale bar = 10 m m |
4. DiscussionThe actual rate of infection on last-year needles could be higher than the current data. Partially diseased needles were casted before they were collected to examine the fungal pustules. In order to obtain accurate result, timing of inventory for diseased needles can be done as early as in September, when disease symptom occurs on last-year needles and have not been releasing basidiospores.
The basidiospore production pink was in the middle of June 1998 (fig. 6). The observation during 1999 and 2000 supposed that. In 2000 the basidiospores begin to occur in the beginning of June. Relative humidity was in long period with 90% in 1998 to compare to 1999. New infection takes place immediately after liberation of basidiospores in optimal conditions.
C. abietis is autoecious rust. In early summer teliospores germinate and give a rise to promycelia which meiosis occurs and on the surface of which sterigmata and basidiospores develop. Upon germination a basidiospore either gives rise to a germ tube or forms an outgrowth that functions as a sterigma, forming a second spore at its tip which this phenomenon is quite common in the laboratory when basidiospores are discharged on to either water or agar medium surfaces (Alexopoulos et al. 1996; Mims and Richard 1990). When a basidiospore lands on current-growth needle, and environmental conditions are conducive, it typically exhibits direct germination through stomata (Grill et al. 1980).
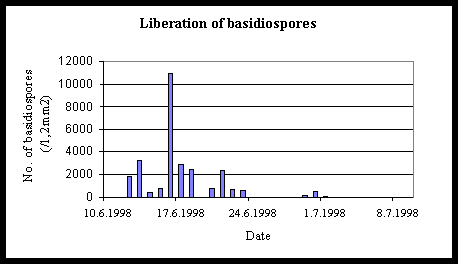
Fig. 6. Numerical record for liberation of basidiospors by the spore trap
5. ReferencesAlexopoulos, C. J., Mims, C. W. and Blackwell, M., 1996. Introductory Mycology. John Wiley and Sons, INC.
600-621.Anonymous, CMI Descriptions of pathogenic fungi and bacteria. No. 576.
Collins, M. A., 1976. Periodicity of spore liberation in Chrysomyxa abietis. Trans. Br. Mycol. Soc. 67 (2): 336-339.
Grill, D.; Lackner, E. and Scharner, M., 1980. Untersuchungen an mit Chrysomyxa abietis befallenen Fichtennadeln.
Phyton (Austria) 19: 71-82.Grill, D.; Pfeifhofer, W. and Esterbauer, H., 1984. Carotenoids in Chrysomyxa abietis infected spruce needles. Eur.
J. For. Path. 14: 296-301.Lehtijärvi, Asko, 1998. Lophodermium piceae and Tryblidiopsis pinastri Two Latent Colonizers of Norway Spruce; Subtopic: Co-occurrence of the ascomycete Lophodermium piceae and the rust fungus Chrysomyxa abietis in Norway Spruce needles. Doctoral thesis. Swedish University of Agricultural Science.
Mims, C. W. and E. A. Richard, 1990. Ultrastructure of secondary spore formation in the rust Gymnosporangium
juniperi-virginianae. Mycologia 82: 236-264.Murray, J. S., 1953. A note on the outbreak of Chrysomyxa abietis Unger in Scotland. Scottish Forestry 7: 52-56.
Peterson, R. S., 1963. Effects of broom rusts on spruce and fir. U. S. Dep. Agric. For. Serv. Res. Pap. INT-7.
Solberg, S. 1996. Local county monitoring plots. Vitality survey 1996. Rapport fra skogforsk. No. 14: 15 pp.
