Serrasalmidae
FIN/SEED/FRUIT EATING PIRANHAS OR CARIBITOS
genus Pristobrycon
Eigenmann 1915
Etymology: Greek, pristis = saw + Greek, bryko = to bite
If you do not want to read this page and just want to access Pristobrycon species list click HERE.
FROM FRANK MAGALLANES
Present study (Freeman, et al.) suggests creating a new genus and placing Pristobrycon striolatus as the type. P. striolatus does not have ectopterygoid (palatine teeth) or a preanal spine. Also considered being placed with P. striolatus is P. careospinus and P. maculipinnis. The other remaining members of this present genus do have the ectopterygoid teeth and preanal spine. The suggestion is these species that do have the preanal spine should be moved into genus Serrasalmus. But that maybe problematic because of the teeth and other structural problems. The use of the tree in that paper may have its own inherent problems. Strongly suggest hobbyists not draw any conclusions based on that work until more work is produced to better evaluate the situation.
Hobbyists and researchers are also reminded that some species have been added and removed several times by historical ichthyologists in this genus. A good example is Serrasalmus eigenmanni. This species has been added and replaced in Pristobrycon and Serrasalmus. It has not been resolved to this day because there are nominal species named in different countries that are problematic.
V. Genus Pristobrycon gen.nov.
Eigenmann: The Serrasalminae and Mylinae, pg. 245-248
Type, *Pygocentrum calmoni; Steindachner.
Intermediate in technical characters between the fierce Rooseveltiella without palatine teeth, and the less blood-thirsty Serrasalmo, with a series of permanent teeth along the palate. They are the least destructive of the piranhas. Head short and deep, the snout short; palate with few or no teeth in the adult, the teeth sometimes, as in aureus, more fully developed in the young; cheeks only partly armed; mouth rather narrow, the upper jaw not very oblique, the lower jaw not very prominent.
Eigenmann placed four species in his new generic ranking. The genus included P. scapularis (Günther), P. aureus (Agassiz), P. calmoni (Steindachner) and P. striolatus (Steindachner).
Key To The Species Of Pristobrycon
Palatine teeth;
a. Sides variously spotted.
6. Margin of caudal pale
c. Depth 1.6-1.8; head 3.12-3.66; D. 15-17; A. 32-34; serrae 27-33; interorbital 2.25-2.5 in the head; profile but little depressed over eye; distance from dorsal to caudal equal to length of head, much longer than dorsal; upper half of sides with numerous very small black spots; .2-.4 of the cheek naked.
scapularis (Giinther). 10. cc. Depth 1.6-1.7; head 3.1-3.75; D. 15-17; A. 32-37; serrae 22-35; interorbital 2.16 in the length of head; profile more depressed over eye; distance from dorsal to caudal longer than head; upper half of sides with larger, more or less prominent spots; .25-.5 of the cheek naked.
aureus (Agassiz). 11.ccc. Depth 1.33; head 3.66; D. 17; A. 40. emarginatus (Schomburgk). 12. 66. Margin of caudal dark. Depth 1.4-1.5; head 3.4-3.66; D. 15-16; A. 32 or 33; serrae 32-33; nearly half of the cheek naked; a small, obscure, humeral spot; small, dark spots on the upper half of body.
calmoni (Steindachner). 13. aa. Numerous dark brown cross-bands dividing below the lateral line into narrow stripes; second suborbital but little higher than eye, its length 1.5 in its height; greatest width of naked area of cheek equal to about one-half the length of the suborbital; head 3.33; depth 1.66; D. 17; A.3ior32; serrae 32; interorbital 2.75; origin of dorsal an orbital diameter nearer to snout than the base of the caudal striolatus (Steindachner). 14. 10. Pristobrycon scapularis (Giinther).
The Species of Pristobrycon
Pristobrycon scapularis, it was noted by Eigenmann; palatines roughened more or less and with tooth-like tubercle.
Pristobrycon aureus, nothing was mentioned about lack or presence of palatine teeth.
Pristobrycon calmoni, palate with one or two teeth.
Pristobrycon striolatus; known only from the types, 180-200mm, long, coming from tributaries of the Rio Pará. No information given.
* Spelling error in the manuscript
Norman (1929), later would consider Eigenmann's work and revised Pristobrycon as a synonym of Serrasalmus. He did recognize differences (i.e., S. scapularis, S. serrulatus, S. striolatus, and S. calmoni) in other species but chose instead to keep the division. With Pristobrycon striolatus, the one species not fully reviewed by Eigenmann in his placement above, Norman found palatines sometimes roughened, but without teeth (page 795, Fishes of the Serrasalmoninae, Norman 1928).
Géry (1972:210) treated Pristobrycon like a subgenus with vague characters to differentiate the species including; high backed, discoid formed, with small scales covering the fleshly part of the dorsal and anal fin. He included other characters pertaining to the size of the mouth not dissimilar from Eigenmann (1915). Géry also disputed the validity of certain new species and decided that only 4 species were valid (i.e., calmoni, striolatus, aureus and serrulatus). These last two included two subspecies; aureus-gymnogenys and serrulatus-scapularis. The rest of the species he considered them synonyms. Géry offered the best yet placement of cataloged species from the Guyana.
Machado-Allison (1982a, 1983, and 1985) discusses the validity of the genera in this subfamily using Phylogenetic analysis concluding that this genus is more closely related to Serrasalmus than with any another group of the subfamily. In the manuscript; Los Peces Caribes de Venezuela (Machado-Allison and Fink, 1996) a new key was proposed for Pristobrycon, pg. 7: 3a. Cabeza robusta con el perfil dorsal convexo. Ancho de la cabeza (12.5-18.9, usualmente 15.5% del largo estándar). Sierras prepélvicas 15-20 (usualmente 17). Dientes ectopterigoideos ausentes en juveniles y adultos. [trans: Robust head, convex dorsal profile. Wide head (12.5-18.9 usually 15.5% of the standard length). Serrae prepélvicas 15-20 (usually 17). Ectopterygoid teeth absent in young and adult.]. serrulatus, P. striolatus, and P. eigenmanni are very similar appearing species. Body shapes are very close and delineating the species is difficult since body spotting may appear similar depending on ontogeny. The ontogeny of the species when juvenile run almost parallel with S. irritans because of the elongated body and the dark 'V' on the base of the caudal peduncle (hypural margin). The problem's I have encountered in distinguishing the species is the body shape. All of these species are discoid and most possess a small humeral spot. In small sizes, they also resemble some Serrasalmus species.
It is presently unknown, if the presence of a humeral spot (or lack of) has anything to do with water conditions that might effect the pigment on the flank or belly coloration. It is known that Pygocentrus nattereri is found in a variety of colors, depending on several factors in relation to geography as is S. rhombeus. It would seem plausible that the same may hold true of these other types of serrasalmin. Another form of Pristobrycon is P. calmoni. The species calmoni lacks the bars and stripes common to Serrasalmus (Pristobrycon) eigenmanni. Three new species of piranha have been discovered, two of them being Pristobrycon. They are P. maculipinnis and P. careospinus. Pristobrycon, with the exception of P. striolatus, forms a clade with Serrasalmus gouldingi that is the sister group to the Serrasalmus-Pygocentrus clade (Guillermo Orti).
HISTORICAL PLACEMENT
| Pygopristis antoni
Fernández-Yépez [A.] 1965:[1], Fig. [Evencias No. 17;Río San José, Estado Guárico, Venezuela. Holotype: MACLPI 65635-A. Paratypes: AFY 60315 (15, ?now 14), 65413 (1); MBUCV-V-12143 [ex AFY 60315] (1), MACLPI 65635-B (3), 65636 (8). Type catalog: Provenzano et al. 1998:15. On first unnumbered page. •Mention -- (Géry 1972:209). •Synonym of Pygopristis denticulatus (Cuvier 1819) -- (Taphorn 1992:309). •Synonym of Pristobrycon striolatus (Steindachner 1908) -- (Machado-Allison 2002:49, Jégu in Reis et al. 2003:188). Current status: Synonym of Pristobrycon striolatus (Steindachner 1908). |
Serrasalmo aureus
Spix [J. B. von] & Agassiz [L.] 1829:72, Pl. 29 [Selecta genera et species piscium quos in itinere per Brasiliam annos MDCCCXVII-MDCCCXX jussu et auspiciis Maximiliani Josephi I.... colleget et pingendso curavit Dr J. B. de Spix....; Equatorial lakes and rivers of Brazil. No types known. •Valid as Serrasalmus aureus Spix & Agassiz 1829 -- (Géry 1972:218). •Valid as Pristobrycon aureus (Spix & Agassiz 1829) -- (Jégu in Reis et al. 2003:188).
Current status: Valid as Pristobrycon aureus (Spix & Agassiz 1829). Serrasalmidae. Distribution: Lower portions of tributaries of lower Amazon River and Guyana rivers: Brazil and Guyana. |
Pristobrycon baratai
Amaral Campos [A.] 1946:218, Fig. 1 [Papéis Avulsos do Departamento de Zoologia (Săo Paulo) v. 7 (no. 17); Rio Tapajós, Santarém, Pará, Brazil. Holotype: MZUSP [ex DZSASP] 3630. Paratypes: MZUSP [ex DZSASP] 3631 (1). Type catalog: Britski 1969:202, Oyakawa 1996:490-491. •Mention -- (Géry 1972:211). •Synonym of Pristobrycon striolatus (Steindachner 1908) -- (Jégu in Reis et al. 2003:188).
Current status: Synonym of Pristobrycon striolatus (Steindachner 1908). |
Pygocentrus bilineatus
Eigenmann [C. H.] 1909:47 [Annals of the Carnegie Museum v. 6 (no. 1); Aruka River, Guyana. Holotype: FMNH 58075 [ex CM 1072]. Paratypes: (4, CM and IU) CAS 57355-56 [ex IU 11756-57] (1, 2); FMNH 58076 (1). Type catalog: Henn 1928:69, Ibarra & Stewart 1987:73. •Synonym of Serrasalmus calmoni Steindachner 1908, but a valid subspecies -- (Géry 1972:220). •Not a Pygocentrus -- (Fink 1993:671). •Synonym of Pristobrycon calmoni (Steindachner 1908) -- (Machado-Allison 2002:49, Jégu in Reis et al. 2003:188).
Current status: Synonym of Pristobrycon calmoni (Steindachner 1908). |
Serrasalmo (Pygocentrus) calmoni
Steindachner [F.] 1908:361 [Anzeiger der Akademie der Wissenschaften in Wien v. 45 (no. 18); Rio Pará, Brazil. Syntypes: (several) NMW (apparently lost) •Valid as Serrasalmus calmoni Steindachner 1908 -- (Géry 1972:220). •Not a Pygocentrus, type lost -- (Fink 1993:671). •Valid as Pristobrycon calmoni (Steindachner 1908) -- (Machado-Allison 2002:49, Jégu in Reis et al. 2003:188).
Current status: Valid as Pristobrycon calmoni (Steindachner 1908). Serrasalmidae. Distribution: Middle and lower Amazon and Orinoco River basins; coastal rivers of Guyana: Brazil, Guyana and Venezuela. |
|
Pristobrycon careospinus
Fink [W. L.] & Machado-Allison [A.] 1992:69, Fig. 20 [Ichthyological Exploration of Freshwaters v. 3 (no. 1); El Loro lagoon, Río Atabapo, 1 kilometer from San Fernando De Atabapo, Estado Amazonas, Venezuela, 4°00'N, 67°50'W. Holotype (unique): MBUCV-V-8145. Type catalog: Provenzano et al. 1998:14. •Valid as Pristobrycon careospinus Fink & Machado-Allison 1992 -- (Machado-Allison 2002:53, Jégu in Reis et al. 2003:188).
Current status: Valid as Pristobrycon careospinus Fink & Machado-Allison 1992. Serrasalmidae. Distribution: Atabapo River in Orinoco River basin: Venezuela. |
Serrasalmus coccogenis
Fowler [H. W.] 1911:428, Fig. 4 [Proceedings of the Academy of Natural Sciences of Philadelphia v. 63; La Pedrita, Cańo Uracoa, Venezuela. Holotype: ANSP 37870. Paratypes: ANSP 37871-75 (5), FMNH 55906 (1). Type catalog: Böhlke 1984:44, Ibarra & Stewart 1987:79. •Synonym of Serrasalmus calmoni (Steindachner 1908), subspecies bilineaetus (Eigenmann 1909) -- (Géry 1972:220). •Synonym of Pristobrycon calmoni (Steindachner 1908) -- (Machado-Allison 2002:49, Jégu in Reis et al. 2003:188).
Current status: Synonym of Pristobrycon calmoni (Steindachner 1908). |
Serrasalmus eigenmanni
Norman [J. R.] 1929:804, Fig. 16 [Proceedings of the General Meetings for Scientific Business of the Zoological Society of London 1928 (pt 4) (no. 30); Rockstone, Guyana. Holotype (unique): BMNH 1911.10.31.496-497 (1 of 2, 120 mm). Norman mentions only one specimen (type of the species) in the original description. •Valid as Pristobrycon eigenmanni (Norman 1929) -- (Jégu & Santos 2001:136). •Valid as Serrasalmus eigenmanni Norman 1929 -- (Géry 1972:224, Machado-Allison 2002:81, Jégu in Reis et al. 2003:190, Machado-Allison et al. 2009:128).
Current status: Valid as Serrasalmus eigenmanni Norman 1929. Serrasalmidae. Distribution: Amazon River basin and northern and eastern Guiana Shield rivers: Brazil, French Guiana, Guyana, Suriname and Venezuela |
Salmo erythrophthalmus
Kner [R.] (ex Natterer) 1860:43 [35] [Denkschriften der Mathematisch-Naturwissenschaftlichen Classe der Kaiserlichen Akademie der Wissenschaften in Wien v. 18; On p. 35 of separate. Not available, name mentioned in passing under Serrasalmo aureus Spix; from Rio Vaupe, Matogrosso, Brazil. •In the synonymy of Pristobrycon aureus (Spix & Agassiz 1829) -- (Jégu in Reis et al. 2003:188).
Current status: Synonym of Pristobrycon aureus (Spix & Agassiz 1829). |
Pygopristis gibbosus
Starks [E. C.] 1913:17, Pl. 3 [Leland Stanford Jr. University Publications, University Series; Market at Pará, Brazil. Holotype: SU 22226. Paratypes: (several) AMNH 3932 (2), SU 22222 (9). Type catalog: Böhlke 1953:26. The type specimen designated in plate 3. •Synonym of Pristobrycon calmoni (Steindachner 1908) -- (Jégu in Reis et al. 2003:188).
Current status: Synonym of Pristobrycon calmoni (Steindachner 1908). |
|
Serrasalmo gymnogenys
Günther [A.] 1864:371 [Catalogue of the fishes in the British Museum. v. 5; Guyana. Syntypes: (3) BMNH 1971.5.10.59 (1) British Guiana [Guyana], 1849.11.8 [or 3].33 (1) Rio Capin [= Capim], Pará State, Brazil. •Synonym of Serrasalmus aureus (Spix & Agassiz 1829), but a valid subspecies -- (Géry 1972:218). •Synonym of Pristobrycon aureus (Spix & Agassiz 1829) -- (Jégu in Reis et al. 2003:188).
Current status: Synonym of Pristobrycon aureus (Spix & Agassiz 1829). |
Pristobrycon maculipinnis
Fink [W. L.] & Machado-Allison [A.] 1992:66, Fig. 19 [Ichthyological Exploration of Freshwaters v. 3 (no. 1); Cańo Doyotomoni, tributary of Río Pamoni, 4 kilometers south of the confluence with Río Casiquiare, Venezuela, 2°48'N, 64°54'W. Holotype: MBUCV-V-16421. Paratypes: MBUCV-V-20803 (1), MCNG 21757 (2), UMMZ 219203 (2). Type catalog: Provenzano et al. 1998:14-15. •Valid as Pristobrycon maculipiinis Fink & Machado-Allison 1992 -- (Machado-Allison 2002:53, Jégu in Reis et al. 2003:188).
Current status: Valid as Pristobrycon maculipinnis Fink & Machado-Allison 1992. Serrasalmidae. Distribution: Blackwater areas in tributary of Casiquiare River, a tributary of Atabapo River, Venezuela. |
Serrasalmo scapularis
Günther [A.] 1864:368 [Catalogue of the fishes in the British Museum. v. 5; Essequibo River, Guyana. Syntypes: BMNH 1864.1.21.34-36 (3), 1971.5.10.58 (1). •Synonym of Serrasalmus serrulatus (Valenciennes 1849), but a valid subspecies -- (Géry 1972:222). •Synonym [of the younger] Pristobrycon striolatus (Steindachner 1908) -- (Jégu in Reis et al. 2003:188). •Perhaps should be considered as valid as Pristobrycon scapularis (Günther 1864).
Current status: Synonym of Pristobrycon striolatus (Steindachner 1908). |
Serrasalmo (Pygocentrus) striolatus
Steindachner [F.] 1908:360 [Anzeiger der Akademie der Wissenschaften in Wien v. 45 (no. 18); Rio Pará, Brazil. Syntypes: NMW 10583-84 (2). •Not a Pygocentrus -- (Fink 1993:671). •Valid as Pristobrycon striolatus (Steindachner 1908) -- (Taphorn 1992:305, Machado-Allison 2002:49, Jégu in Reis et al. 2003:188). •Valid as Serrasalmus striolatus Steindachner 1908 -- (Géry 1972:218, Planquette et al. 1996:380, Camargo & Isaac 2001:139).
Current status: Valid as Pristobrycon striolatus (Steindachner 1908). Serrasalmidae. Distribution: Amazon and Orinoco River basins and northern and eastern Guiana Shield rivers: Brazil, French Guiana, Guyana, Suriname and Venezuela. |
genus Pristobrycon
Eigenmann [C. H.] 1915:245 Masc. Serrasalmo (Pygocentrus) calmoni Steindachner 1908. Type by original designation. •Synonym of Serrasalmus Lacepčde 1803, but a valid subgenus Pristobrycon -- (Géry 1972:210, Géry 1976:52, Géry 1977:278). •Valid as Pristobrycon Eigenmann 1915 -- (Taphorn 1992:305, Machado-Allison 2002:45, Jégu in Reis et al. 2003:188, Freeman et al. 2007:2 et seq., Mirande 2009:6 in Serrasalmidae, Mirande 2010:471, Hubert & Renno 2010:33.
Current status: Pristobrycon Eigenmann 1915. Serrasalmidae. |
CURRENT STANDING
ABSTRACT - Freeman, B., Nico, L. G., Ostentoski, M. Jelks, H. J. & Collins, T. M. 2007. Molecular systemics of Serrasalmidae: Deciphering the identities of piranha species and unraveling their evolutionary histories. Zootaxa 1484: 1 -38:
Piranhas and their relatives have proven to be a challenging group from a systematic perspective, with difficulties in identification of species, linking of juveniles to adults, diagnosis of genera, and recognition of higher-level clades. In this study we add new molecular data consisting of three mitochondrial regions for museum vouchered and photo-documentedrepresentatives of the Serrasalmidae. These are combined with existing serrasalmid sequences in GenBank to address species and higher-level questions within the piranhas using parsimony and Bayesian methods. We found robust support for the monophyly of Serrasalmus manueli, but not for Serrasalmus gouldingi when GenBank specimens identified as S. gouldingi were included in the analysis. “Serrasalmus gouldingi” sequences in GenBank may, however, be misidentified. Linking of juveniles to adults of the same species was greatly facilitated by the addition of sequence data. Based on our sampling and identifications, our data robustly reject the monophyly of the genera Serrasalmus and Pristobrycon. We found evidence for a well-supported clade comprised of Serrasalmus, Pygocentrus, and Pristobrycon (in part).
This clade was robustly supported in separate and combined analyses of gene regions, and was also supported by a unique molecular character, the loss of a tandem repeat in the control region. Analysis of specimens and a literature review suggest this clade is also characterized by the presence of a pre-anal spine and ectopterygoid teeth. A persistent polytomy at the base of this clade was dated using an independent calibration as 1.8 million years old, corresponding to the beginning of the Pleistocene Epoch, and suggesting an origin for this clade more recent than dates cited in the recent literature. The sister group to this clade is also robustly supported, and consists of Catoprion, Pygopristis, and Pristobrycon striolatus. If the term piranha is to refer to a monophyletic clade, it should be restricted to Serrasalmus, Pygocentrus, and Pristobrycon (in part), or expanded to include these taxa plus Pygopristis, Catoprion, and Pristobrycon striolatus. Accepted by Gilbert: 31 Mar. 2007; published: 28 May 2007
|
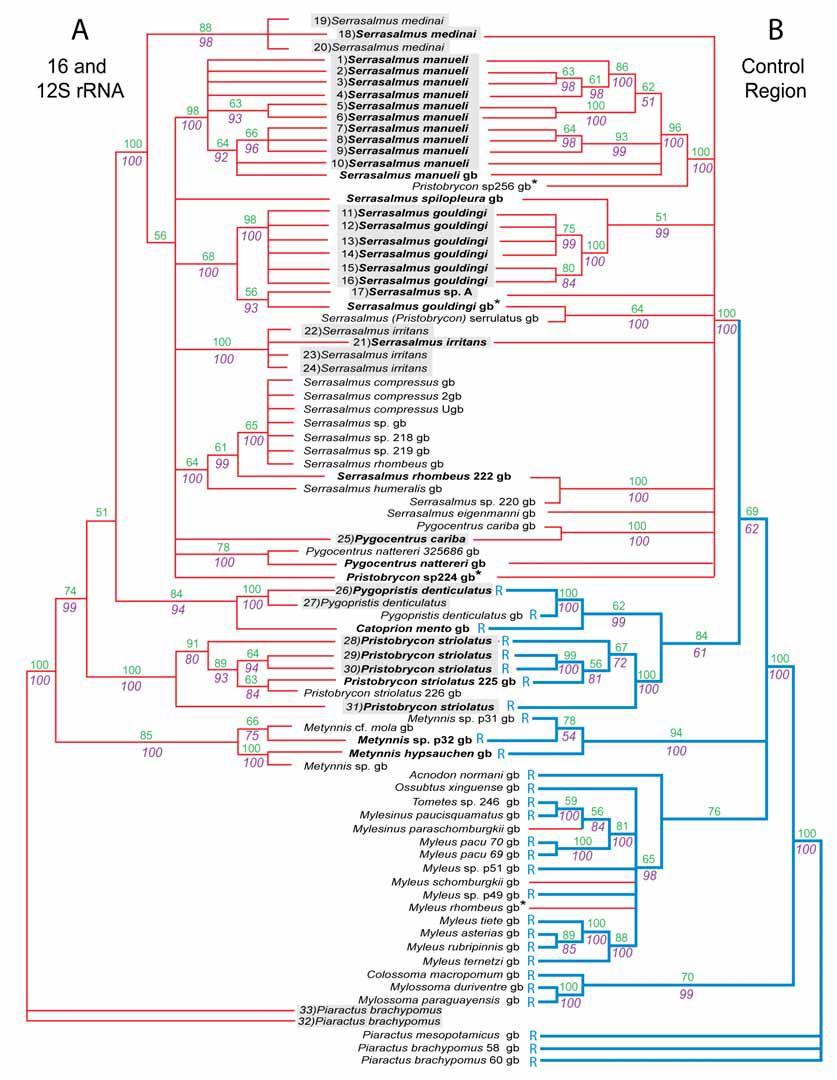
FIGURE 7.Phylogenetic trees of serrasalmids inferred from ribosomal (A) and control region (B) data sets, both including original material and GenBank sequences. Parsimony bootstrap percentages are shown above branch nodes, while proportions (>50%) of trees possessing a given clade in Bayesian posterior distributions are shown below. Taxa shared by both trees are in bold font. Specimen sequences from original material appear in shadow boxes, preceding numbers (1-33) correspond to numbered specimens and information presented in Table 1 and elsewhere. GenBank sequences are followed by gb. Taxa with VNTR in control region marked by an “R” and reconstruction of presence of VNTR shown in blue. GenBank (gb) species with asterisk (*) indicate taxa of which we question the identification. |
FIGURE 8. Phylogram of combined ribosomal and control region sequences. Analysis includes specimens appearing in bold font in Figure 7. Proportion of trees from posterior distribution possessing a given clade below branch, parsimony bootstrap proportions (>50%) above branch. Specimen sequences from original material appear in shadow boxes, associated number in parentheses (1–33) correspond to numbered specimens and information presented in Table 1 and elsewhere. GenBank sequences are followed by gb. Arrow marks branches with lengths that were not significantly different from zero. GenBank (gb) species with asterisk (*) indicate taxa of which we question the identification |
CONCLUSIONS
Piranha systematics are undergoing major revision and much remains to be done. For example, a functional key to identify many juvenile and adult piranhas is lacking. In spite of the hurdles remaining, we are confident that continued and more detailed genetic analyses combined with further scrutiny of morphological characters will ultimately produce a clear picture of piranha and serrasalmid phylogenetic relationships. Progress will require inclusion of other Serrasalmus and all or most of the remaining Pristobrycon species. Of particular importance relative to the piranha clade is the need to determine if genetic analysis consistently separates all Pristobrycon species with pre-anal spines from those without and if, as has so far been shown with P. striolatus, those without a pre-anal spine are separate from the clade consisting of piranhas with pre-anal spines. (Freeman et al.)
Given the continued confusion and complexity, the work remaining is still substantial. Future studies of serrasalmid phylogeny should include: 1) combined genetic data, including additional unlinked loci and anatomical analysis of new material and reanalysis of older specimens, and 2) careful documentation of specimens examined, including vouchering of all material and establishment of photographic archives of specimens studied. (Freeman et al).
2012 CURRENT SPECIES CLICK HERE.
REFERENCES
Eigenmann, C. H. 1915 (Mar.) The Serrasalminae and Mylinae. Ann. Carnegie Mus. v. 9 (nos. 3-4): 226-272, Pls. 44-58.
Géry, J. 1972 (19 Dec.) Poissons characoďdes des Guyanes. I. Généralités. II. Famille des Serrasalmidae. Zool. Verh. (Leiden) No. 122: 1-250, Pls. 1-16.
Géry, J. 1976 (18 Mar.) Les genres de Serrasalmidae (Pisces, Characoidei). Bull. Zool. Mus. Univ. Amst. v. 5 (no. 6): 47-54.
Géry, J. 1977 Characoids of the world. T. F. H. Publications, Neptune City, New Jersey. Characoids World: 1-672.
MACHADO-ALLISON, Antonio and FINK, William., 1996 - Los Peces Caribes de Venezuela, Diagnosis, Claves, Aspectos Ecologicos Y Evolutivos, pg. 29-31.
FINK, William and MACHADO-ALLISON, Antonio., Three new species of piranhas from Brazil and Venezuela. Ichthyological Explorations of Freshwaters, 3(1):55-71. 1992.
Taphorn, D. C. 1992 The characiform fishes of the Apure River drainage, Venezuela. Biollania Edición Especial - No. 4. Monografias Cientificas del Museo de Ciencias Naturales, UNELLEZ -- Guanara, estado Portuguesa, Venezuela. 1-537.
Freeman, B., Nico, L. G., Ostentoski, M. Jelks, H. J. & Collins, T. M. 2007. Molecular systemics of Serrasalmidae: Deciphering the identities of piranha species and unraveling their evolutionary histories. Zootaxa 1484: 1 -38
California Academy of Sciences. On-line: http://research.calacademy.org
USE THIS LINK TO RETURN TO RESEARCH PAGE.
The OPEFE web site and its contents; is disclaimed for purposes of Zoological Nomenclature in accordance with the International Code of Zoological Nomenclature, Fourth Edition, Article 8.3 and 8.4. No new names or nomenclature changes are available from statements at this web site.
Copyright© 1994-2016 Oregon Piranha Exotic Fish Exhibit (The OPEFE fish exhibit is permanently CLOSED as of 2000) Sutherlin, Oregon. Information posted on this web site is archival data on fish scientific classifications and other information. DISCLAIMER: The copyrighted material may not be used for any purpose other than private study, scholarship or research. Cited information requires credit and this link www.opefe.com. All rights reserved. All images shown (unless otherwise noted) is property of OPEFE.
UPDATED: 12/06/2015