|
A. Beneficial Fungal Parasites 1. Fungi as Biocontrol Agents
|
|
|
Two of the
main areas in which parasitic fungi are beneficial to man are in the use
of fungi to control pests and in the prevention of herbivory in plants. Biocontrol
is the reduction of the amount of inoculum or disease producing activity
of a pathogen accomplished by or through one or more organisms other than
man. Biocontrol uses natural or modified organisms, genes or gene products
to reduce the effects of undesirable pests and to favor desirable
organisms such as crops, trees, animals and beneficial insects and
microorganisms.
The prime tactics used in biocontrol involve (1) modifying cultural
practices to favor antagonistic plant pathogens, (2) introducing
antagonists of plant pathogens into the environment or onto the plant, and
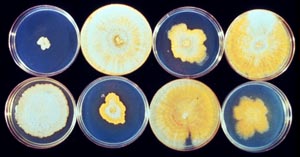
(3) inoculate the plant with incompatible or hypovirulent organisms. a. Fungi to Control Plant Diseases: Two yeasts have been pinpointed that produce killer toxins that block the growth of other yeasts (Ag. Res., June 1992, USDA). The yeast, Pichia inositovora and P. acaciae, have been shown to produce antifungal cytotoxic proteins that will inhibit the growth of a number of other yeasts. This phenomenon has been put to work where toxins of Kluyveromyces lactis fights against unwanted yeasts that may contaminate fermenting “sake” or rice wine. Species of Trichoderma are used to control root diseases of many crops, stem blight of peanuts, dry bubble on mushrooms, and silverleaf of plums, Verticillium to control cotton wilt, Sphaerellopsis to control rust diseases on a number of plants, and several others. These belong to an array of mycofungicides. Prime examples of some are given below: Chestnut blight, caused by the perithecial fungus Cryphonectria parasitica, entered the U.S. on European chestnut timber brought into lumber mills in New England. The fungus was extremely virulent to North American chestnuts that had never been exposed to the fungus. Chestnuts were then the dominant hardwood lumber tree in this country. The pathogen spread first into New York, being first detected on chestnuts at the New York Botanical Garden in 1904. It soon spread throughout the range of the American chestnut into the southern Smoky Mountains where by 1935 essentially all trees had been killed (Fig. 10-1).
Fig. 10-1. A chestnut tree that was killed in the 1920s by Cryphonectria parasitica, the chestnut blight fungus. In subsequent years, efforts to find a control for this fungus were fruitless. A biological control imported from Europe in 1972 allows us to keep American chestnut trees alive. Strains of C. parasitica from Europe were shown to hypovirulent, i. e. lower virulence that the native strains. It was initially shown that the hypovirulent strain, once inoculated onto a tree diseased with the native strain, would dominate and suppress the virulence of the native strain, thus allowing the American chestnut trees to survive and grow (Fig. 10-2).
Fig. 10-02. Canker on a young chestnut tree caused by the chestnut blight fungus. Several different hypovirulent strains of C. parasitica were tested for their ability to dominate the virulent strain (Fig. 10-3).
Fig. 10-3. Several hypovirulent strains of Cryphonectria parasitica. (Photo provided by Sandra Anagnostakis) The most promising ones were inoculated into existing infected chestnut trees (Fig. 10-4).
Fig. 10-4. Inoculating chestnut seedlings with a hypovirulent strain of the chestnut blight fungus. (Photo provided by Sandra Anagnostakis) It
was later shown that the hypovirulence in certain strains was the results
of a viral infection of the fungus (Anagnostakis & Hillman, 1992.
Arnoldia 52:3-9). Root rot of Conifers, caused by Heterobasidium annosum, is a devastating disease of southeastern yellow pines. The disease is spread largely through infestation of freshly cut stumps of pines, the spread of the fungus into the decaying stump and into the roots. Through natural root grafts, the fungus may be spread throughout a large area (Fig. 10-5).
Fig. 10-5. Root rot of pine caused by the bracket fungus Heterobasidium annosum. This
disease may be controlled by treating the surface of cut pine stumps with
a spore suspension of Phlebia
gigantea. Colonization of stump surfaces by P.
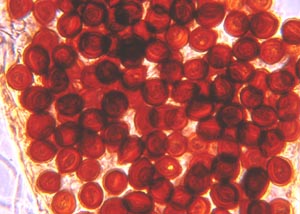
gigantea prevents subsequent colonization by H. annosum. Post-harvest rot of citrus fruit is caused by species of Penicillium, commonly P. chrysogenum. Biocontrol of this problem can be accomplished by applying the yeast, Pichia guillermondii to fruit after harvest but before storage or shipping. Pichia inoculated fruits are shown on the right (Fig. 10-6) and uninoculated on the left.
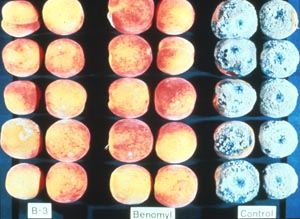
Fig. 10-6. Fruit rot of citrus showing the effectiveness of biocontrol with Pichia guillermondii (U.S.-7). Brown rot of peach or other stone fruits such as plums, apricots, and cherries is caused by the discomycete Monilinia fruticola and can be a major problem during rainy seasons, often causing total crop losses. Benomyl is the common fungicide used to control brown rot. The disease, however, may be controlled by inoculating the fruits with the bacterium Bacillus subtilis after harvest but before storage or shipping. This bacterium produces an antifungal metabolite that is toxic to M. fruticola. Fig. 10-7 compares the effectiveness of biocontrol to that of benomyl.
Fig. 10-7. Biocontrol of brown rot of peach comparing Bacillus subtilus with the fungicide Benomyl. Damping-off affects seeds, seedlings, and succulent roots of many plants. Losses can be great in moist soils when temperatures are low. Damping off of cotton is caused be the Oomycete Pythium ultimum. A biocontrol tactic used is to treat soil with the fungus Gliocladium virens before planting the cotton seed. The production of the antifungal compound, gliovirin, plays an important role in the control of damping-off. This fungus has also been used in the control of another important root pathogen, Rhizoctonia solani. Verticillium wilt of potato is caused by Verticillium dahliae. A biocontrol practice used on this disease is to modify cultural practices to stimulate antagonists or competitors of. V. dahliae. Crop rotation and solarization helps the soil to become suppressive to Verticillium. b. Biocontrol of Insects: The knowledge of entomogenous fungi dates back for several centuries (McCoy et al. 1988. in Ignoffo, Handbook of Natural Pesticides. CRC Press, Boca Raton, FL). Numerous groups of entomogenous fungi were described during the 19th century. Pasteur (1874. C.R. Acad. Sci. 79:1233), however, was one of the first to suggest that microorganisms could be used to control insect pests. Extensive research on the use of Beauvaria to control chinch bugs in Kansas had mixed success (Snow, 1896. Ann. Rep. Kan. Exp. Sta. 5:7). One of the earliest successes in biocontrol was the use of Aschersonia aleyrodes to control citrus white flies in Florida (Berger, 1921. Bull. State Pl. Bd. 5:141). More than three decades ago, Rachael Carson's Silent Springs pointed out the dangerous effects of a number of our most effective pesticides. This eventually led to the prohibition of the use of chlorinated hydrocarbons as insecticides. Her book sparked considerable interest in finding alternative ways of controlling insects. By the 1960s there were a number of attempts for the commercial production of fungi for biological control. Close to 750 species of entomogenous fungi had been identified by the 1980s (Microbial Biocontrol Agents. Nat. Acad. Sci. Washington, DC, 1979). Almost all of the major groups of fungi have entomogenous species, many of them have been used as mycoinsecticides. (I thank Dr. Clay McCoy, Citrus REC, Lake Alfred, FL for the use of some images of entomogenous fungi.) Quite a number of insects can be controlled with fungi; these include the cabbage loopers (Fig. 10-8) in which the body cavity becomes overwhelmed with spores (Fig. 10-9);
Fig. 10-8. The control of cabbage loopers with the fungus Noumorea rileyi.
Fig. 10-9. Insect cavity filled with fungal spores. species of Cordyceps that infects the larvae of many beetles and moths (Fig. 10-10), even those deeply embedded in the soil (Fig. 10-11);
Fig. 10-10. An insect larva infected with a species of Cordyceps.
Fig. 10-11. Insect larvae with several perithecial stroma. species of Zoophora on flies; Stilbella on weevils (Fig. 10-12),
Fig. 10-12. A weevil highly infected with Stilbella. Hirsutella on the larva of a citrus mite, Paecilomyces on beetle larvae (Fig. 10-13), and species of Beauvaria (Fig. 10-14) that will infect a large number of insects.
Fig. 10-13. Colonies of Paecilomyces variotii growing out of a beetle larva.
Fig. 10-14. Beauvaria bassiana infecting a weevils. In Florida, species of Aschersonia commonly infects citrus white flies and Noumorea on soybean looper (Fig. 10-15).
Fig. 10-15. Noumorea rileyi infecting a soybean looper. Species of Metarrhizium infects a number of insects (Fig. 10-16), forming long chains of spores (Fig. 10-17); a feature that has enabled its use in novel roach traps using the fungus rather than chemicals (Fig. 10-18).
Fig. 10-16. A beetle larva infected with a species of Metarrhizium.
Fig. 10-17. Chains of conidia characteristic of Metarrhizium.
Fig. 10-18. Roach chambers that use Metarrhizium instead of chemicals for control. The
use of a fungus in roach traps is superior to chemical use because
chemicals will kill only the insects that enter the chamber; whereas,
insects that become infected with Metarrhizium will carry the
fungus to their hiding places and infect their neighbors. Isn’t that
neat?! One of the most puzzling problems in insect control has been the control of mosquitoes. Mosquitoes have long been of concern to people because their bites are painful and they transmit some of our most important diseases. Dr. John Couch and his students and colleagues at the University of North Carolina spent several years on studies of species of the aquatic fungus Coelomomyces that infects mosquitoes. After exhaustive research projects, they were never able to reinfect healthy mosquitoes with zoospores released from infected ones. It was not until the studies of Dr. Howard Whisler at the University of Washington that the answer came. Whisler found that Coelomomyces was heteroecious, i.e. a species that requires two hosts on which to complete its life cycle. The primary host of this fungus was mosquitoes and the alternate host was a small copepod. Zoospores released from the copepod fused to form a flagellated zygote that was capable of infecting mosquitoes (Fig. 10-19).
|
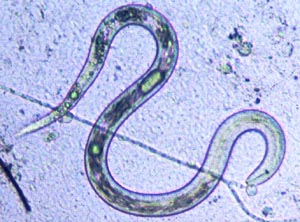
c. Biocontrol of Nematodes: Nematodes (Fig. 10-20) are small, almost microscopic, needle-shaped worms that live in marine and fresh water systems, in soil, and inside of plants and animals. Plant pathogenic nematodes cause diseases of a large number of crop plants and are costly to control.
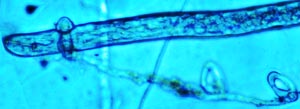
Fig. 10-20. The nematophagus fungus Arthrobotrys candida. Thousands of dollars of crop losses occur annually. Most control procedures involve chemicals, many of which are detrimental to our environment. The use of fungi for biocontrol of nematodes, myconematocide, has been limited. Some success has been experienced, however, with infesting seedlings or soil with species of a nematophagus fungus, Dactyella. Species of Dactyella and Arthrobotrys are well known as nematode trapping fungi. They have peculiar nets (Fig. 10-21), constricting rings (Fig. 10-22), knobs (Fig. 10-20) that trap nematodes.
Fig. 10-21. Formation of a net-like trap by Arthrobotrys oligospora.
Fig. 10-22. Arthrobotrys brocophaga developing constricting ring traps. Once the hyphae trap a nematode, it will invade the body cavity, resulting in death. Aquatic nematodes are susceptible to attack by species of the aquatic oomycete Lagenidium. d. Biocontrol of Weeds and Noxious Plants: There are about 30,000 species of plants that are
considered to be weeds and are directly or indirectly noxious to humans
and their domesticated animals. About 1,600 of these can cause serious
crop losses, with many crops having several competing weeds and costing
close to $14 billion dollars in losses and control of these pests. In
order to control weeds and prevent these losses, agriculturalists have
turned increasingly to the use of herbicides. As you are well aware
through newspaper and TV reports, many of the herbicides have gotten
into our drinking water and lakes and have the potential of causing
serious health problems. Several large cities in the Midwest have
recently detected alarming levels of herbicides and other pesticides in
their water systems. Mycoherbicides
have advantages over chemical herbicides in that they can be more
host-specific, preparation costs will be less expensive, and human
health hazards can be eliminated. Biological control of weeds is a
positive avenue to approach this problem. There have been some exciting
breakthroughs in recent years. Mycogen Co. in San Diego, CA plans to
market a number of mycoherbicides including Alternaria cassiae to control sicklepod
and Fusarium to control the velvet
leaf weed, both major weed problems in soybean fields. It will take
the cooperation of professionals in plant pathology, weed science, and
fermentation science to bring success in biocontrol of weeds Numerous fungi have been tested for weed control. Skeleton weed that invades crop and pasture lands (Fig. 11-1) has been shown, under greenhouse conditions and field tests (Fig. 11-2), to be controlled by species of the rust fungus Puccinia.
Fig. 11-1. Skeleton weed is a major pest in range land.
Fig. 11-2. The use of a species of Puccinia in the control of skeleton weed in range land. Another species of rust has also been tested for the control of thistles (Fig. 11-3).
Fig. 11-3. Species of Puccinia are used in the biocontrol of thistles.
A close-up of a thistle leaf shows yellowing and death caused by the
rust and recently inoculated
leaves bear numerous rust lesions (Fig.
11-4).
Fig. 11-4. A thistle leaf showing prolific growth of the rust Puccinia. Strangler vine: Phytophthora palmivora has been used to control milkweed or strangler vine, a major problem on citrus in south Florida (Fig. 11-5).
Fig. 11-5. Severe outbreak of strangler vine on citrus in south Florida. The mycoherbicide “Divine”
by Abbott Labs incorporates P. palmivora (Fig.
11-6)
in the biocontrol of this
weed pest.
Fig. 11-6. Research on the use of Phytophthora palmivora to control strangler vine. Jointvetch: “Collego” (Fig. 11-7) is a relatively new mycoherbicide that utilizes Colletotrichum gloeosporoides to control jointvetch where in rice fields, chemical control presents a real problem.
Fig. 11-7. A label from a container of Collego. Several preparations of collego are available. Jointvetch can be a major problem to rice farmers (Fig. 11-8) because in the harvesting of rice, the jointvetch seeds, that are similar in size, contaminate the rice and lowers its market value (Fig. 11-9).
Fig. 11-8. A rice field heavily infested with jointvetch (white arrow heads).
Fig. 11-9. Black jointvetch seeds can heavily contaminate rice grains at harvest. Special culturing techniques are used to produce large volumes of inoculum (Fig. 11-10) before aerial application. Jointvetch plants become infected by Colletotrichum (Fig. 11-11), resulting in defoliation and death.
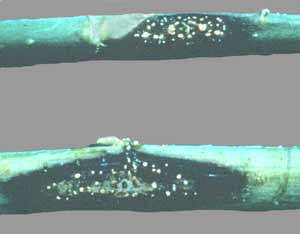
Fig. 11-10. Production of inoculum of Colletotrichum gloeosporoides.
Fig. 11-11. Jointvetch stems infected with Colletotrichum gloeosporoides. Sicklepod, another member of the pea family, has long pods (Fig. 11-12) and is a problem to a number of crops.
Fig. 11-12. Sicklepod is a weed problem in many crops. Alternaria cassiae has been found to be pathogenic to sicklepod and in experimental tests it was shown to kill sprayed plants, while unsprayed plants grew vigorously. Similar results were found in field tests (Fig. 11-13).
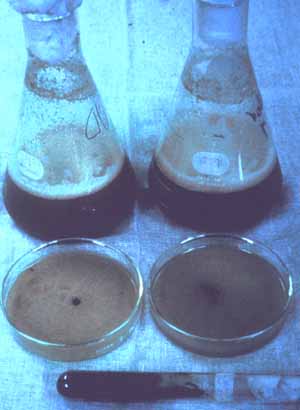
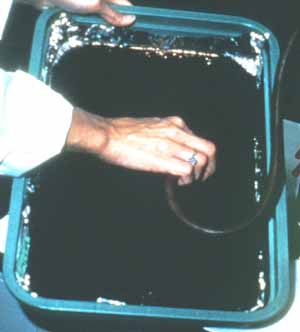
Fig. 11-13. Biocontrol of sicklepod in a soybean field. Inoculum of Alternaria is prepared in large fermentation tanks, pelletized for field inoculations, or grown on large trays of solid media (Fig. 11-14) in which after sporulation, millions of spores are vacuumed (Fig. 11-15) and used for spore inoculations.
Fig. 11-14. Mass production of inoculum of Alternaria cassiae.
Fig. 11-15. Vacuum harvesting of Alternaria spores. Waterhyacinth was first introduced into Florida in the 1890s. Within the past century, it has spread over almost ½ million acres of waterways. It is a beautiful plant (Fig. 11-16) and was very likely spread by people who put them in the gold-fish ponds, fountains, and other bodies of water from which it escaped.
Fig. 11-16. Luxuriant growth of waterhyacinth in freshwater lakes and streams. In 1971, a sharp decline of waterhyacinth was noted in the now famous Rodman Reservoir. It was soon determined that it was highly infected with a species of Cercospora (Fig. 11-17), later described C. rodmanii (Conway, 1976, Can. J. Bot. 54:1179-1183), renamed C. piaropi.
Fig. 11-17. A waterhyacinth leaf heavily infected with Cercospora piaropi. Tests were done on waterhyacinth in Lake Alice at the University of Florida where the results were very promising (Fig. 11-18).
Fig. 11-18. Effective control of waterhyacinth with Cercospora. While good biocontrol was achieved, difficulties were
experienced in the application of inoculum and the survival ability of
waterhyacinth. Hydrilla
perhaps causes the most
problems to fishermen (or should that be fisherpersons). While fungi
that are pathogenic to hydrilla have been discovered, attempts to
control hydrilla present a number of problems. The major one is, how to
apply inoculum to a submerged plant. While fair control was achieved
under laboratory conditions,
it could not be repeated in the field. A
very active research program on the biocontrol of weeds is conducted at
the University of Florida under the supervision of Dr. Charudattan. Some
of the success stories include the use of a new fungus Phomopsis
amaranthicola to control of pigweed; Dactylaria
higginsii to control purple
sedge; Dreschlera/Exerohilum
cocktail to control various
grasses; Bipolaris saccharii
to control cogon grass; and Uromycicola tepperianium to control invading locust trees. (Slides on biological control of weeds were provided by R. Charudattan, Univ. of Florida and Gerald Van Dyke, North Carolina State Univ.) Marijuana: In the summer of 1999, biocontrol became a household word when TV and newspaper releases appeared that told about the use of a pathological race of Fusarium oxysporum that was specific to marijuana and was currently being tested to control this drug plant. Issues were soon raised as to whether biocontrol was safe, would this fungus spread to agriculturally important crops, could the species mutate and form more devastating forms, and what if...? Interestingly, no one has raised similar issues about controlling nutgrass, coffeeweed, sicklepod, and an array of noxious weeds with Fusarium and related fungi.
|