II. BIOLOGÍA CELULAR. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Introducción Un vasto período evolutivo separó la aparición de las primeras moléculas biológicas de las primeras estructuras que podían considerarse vivientes. La evolución permaneció muy restringida hasta que complejos agregados de éstas moléculas se separaron de su medio. En ocasiones, moléculas parecidas a proteínas forman en el agua, de manera espontánea, esferas vacías, llamadas microesferas. Tal vez, hace varios miles de millones de años, algunas de éstas microesferas atraparon en su interior otras moléculas biológicas que formaban una especie de precélula. Las primeras colecciones de moléculas que pudieran considerarse con vida, seguramente poseían una cubierta delgada, una simple membrana celular, que separaba la vida de la ausencia de vida. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
II.1. Célula – definiciones |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
En el mundo vivo la unidad fundamental es la célula, y los procesos vitales ocurren dentro de ella. Las células pueden romperse triturándolas o extraer partículas que efectúan algunos de los procesos vitales, pero es sólo dentro de una unidad como la célula, que pueden ocurrir los procesos del sistema vivo. Una célula puede definirse como la unidad regular de la actividad biológica, rodeada por una membrana y capaz de reproducirse independientemente de cualquier otro sistema vivo. Los organismos vivos, grandes y pequeños, vegetal o animal, pez o ave, ser humano y microbio, están compuestos de células. Las células son básicamente similares entre sí y tienen muchas características estructurales en común. Una de las maravillas de la naturaleza consiste en que una interminable variedad de organismos vivos están construidos o dependan de unidades básicamente semejantes. ¿Cómo se puede reconocer una célula cuando se le ve? ¿Cuáles son las características universales de las células? ¿Qué se quiere decir exactamente cuando se afirma que la célula es la unidad en la que se basan todos los sistemas vivos? Al responder primero a la última pregunta se hallará la respuesta para las dos primeras. Es más fácil comprender el concepto de célula, comparando los organismos vivos con los edificios; las habitaciones de los edificios son análogas a las células de los organismos. Tanto las habitaciones como las células tienen límites, con salidas y entradas: por ejemplo, las habitaciones tienen paredes, pisos, techos, puertas y ventanas, y las células tienen paredes o membranas con poros de varios tamaños. Tanto las habitaciones como las células, se dan en una gran variedad de tamaños, formas, contenidos; cada tipo de habitación o de célula tiene su uso, función o especialidad particular. Los edificios pueden constar de una habitación o de muchas. De la misma manera, los organismos pueden constar de una sola célula, en cuyo caso se denominan unicelulares, o de muchas, esto es, multicelulares. Mediante la combinación de habitaciones de varios tipos, puede construirse una gran variedad de edificios: departamentales, escuelas, oficinas y otros más. Similarmente, los diferentes organismos están construidos por diversos tipos de células. Así como no hay edificios sin habitaciones, tampoco hay vida sin células. Célula. Unidad mínima de un organismo capaz de actuar de manera autónoma. Todos los organismos vivos están formados por células, y en general se acepta que ningún organismo es un ser vivo si no consta al menos de una célula. Algunos organismos microscópicos, como bacterias y protozoos, son células únicas, mientras que los animales y plantas están formados por muchos millones de células organizadas en tejidos y órganos. Célula. Es la unidad de vida más pequeña, formada, por lo menos, de una membrana exterior que rodea a un medio acuoso que contiene moléculas orgánicas, incluido el material genético compuesto de DNA. Célula. Unidad microscópica de estructura y función que comprenden los cuerpos de plantas y animales. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
II.2. Teoría celular |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
“La causa de la nutrición y el crecimiento no reside en el organismo como un todo sino en las células, que son sus partes elementales e individuales.” Theodor Schwann (1810-1882)
Rudolph Virchow (1821-1902) El conocimiento humano de la naturaleza celular de la vida fue lento. En 1665, un científico e inventor inglés llamado Robert Hooke (1635-1703) informó de algunas observaciones con un microscopio primitivo; (véase fig. 10). Colocó en su instrumento un “pedazo de corcho muy delgado” y vio “una gran cantidad de pequeñas celdillas”. Hooke llamó a éstas celdillas “células” porque le recordaban las pequeñas habitaciones o celdas, ocupadas por los monjes. El corcho proviene de la corteza externa seca del alcornoque. Hooke escribió que en éste árbol vivo y en otras plantas, “dichas celdillas están llenas de jugos”. Hooke no dijo lo que eran éstas células y como se relacionaban con la vida de todas las plantas.
En 1673, el inventor holandés, Anton Van Leeuwenhoek (1632-1723) dio a conocer a la Sociedad Real Británica sus observaciones acerca de los eritrocitos, espermatozoides y de una gran cantidad de “animáculos” microscópicos contenidos en el agua de los charcos. Pasó más de un siglo antes de que los biólogos comenzaran a entender la importancia de las células en la vida en la tierra. Los microscopistas primero se dieron cuenta de que muchas plantas estaban formadas completamente por células. La pared gruesa que rodea a todas las células de las plantas hizo que éstas observaciones fueran fáciles. Sin embargo, las células animales fueron descubiertas hasta 1830, cuando el zoólogo alemán Theodor Schwann (1810-1882) vio que el cartílago contiene células que “semejan exactamente a las células de las plantas”. En 1839, después de estudiar las células durante años, Schwann publicó su teoría, llamando células a las partes elementales, tanto de plantas como de animales. A mediados de 1800, un botánico alemán llamado Matthias Schleiden (1804-1881), tuvo una visión científica mas refinada de las células al escribir “... es fácil percibir que los procesos vitales de las células individuales deben formar los fundamentos básicos absolutamente indispensables de la vida”. En pocos años, varios microscopistas habían observado que las células vivas podían crecer y dividirse en células más pequeñas. A finales de 1850, el patólogo austríaco, Rudolph Virchow (1821-1902) escribió: “cada animal es la suma de sus unidades vitales, cada una de las cuales contienen todas las características de la vida”. Es más, Virchow predijo: “todas las células provienen de otras células”. Los tres principios de la teoría celular moderna evolucionaron directamente de los enunciados de Virchow. Postulados de la teoría celular: 1. Unidad anatómica. Cada organismo vivo está formado por
una o más células.
A finales del siglo XIX, se aceptó también que la célula
es la base para comprender las enfermedades, es decir, cuando la gente
enferma es porque sus células están enfermas. Hasta mediados
del siglo XX, la patología (el estudio de las enfermedades) se
apoyó en forma casi exclusiva en el enfoque celular (citológico).
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
c) Actividad de aprendizaje – Teoría celular |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Para comprender la importancia de la formulación de la teoría celular en el contexto de la época convendría leer la novela Cazadores de microbios de Paul de Kruif, publicado por varias editoriales, entre ellas Editores Mexicanos Unidos en 1976. En esta obra se hace referencia al descubrimiento del mundo microscópico con las anécdotas y planteamientos que rodearon las primeras observaciones. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
II.3. Célula procariótica y eucariótica |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Todos los seres vivos se ubican en los siguientes cinco
reinos: Monera, Protista, Fungi, Plantae y Animalia. Es muy posible que los primeros organismos vivos del planeta se parecieran a las Moneras. Los reinos Protista, Fungi, Plantae y Animalia tienen células eucarióticas. A Eduard Chalton, biólogo francés se le atribuyen los conceptos de procarionte y eucarionte. Las células procarióticas (del griego “antes del núcleo”) son por lo general muy pequeñas (de menos de 5 micrómetros de largo), con una estructura interna relativamente simple, casi todas las células procarióticas están rodeadas por una pared celular relativamente dura. Las substancias que forman la pared celular son secretadas por la célula misma. Este tipo de células está representado por las bacterias; (véase fig. 11). Muchas de ellas viven en medios líquidos, desde charcas hasta el torrente circulatorio humano, desde donde el agua tiende a penetrar a la célula bacteriana. Sin una pared celular y sin mecanismos especializados de energía intensa para expulsar el agua, la entrada de agua del medio haría que la célula se hinchara y explotara; de hecho, la penicilina y algunos otros antibióticos combaten las infecciones bacterianas, al inhibir la síntesis de la pared celular y al ocasionar la ruptura de la bacteria. En algunas bacterias, la pared celular tiene una capa de polisacáridos que impide que los leucocitos (que forman parte del sistema inmune del cuerpo humano) ingieran a la bacteria. Aunque realiza una función de sostén importante, la pared celular y su capa de polisacáridos son bastante porosas. El movimiento de materiales hacia el interior y exterior de una célula procariótica es regulado por la membrana plasmática que se encuentra al interior de la pared celular. El citoplasma de la mayor parte de las células procarióticas es en apariencia relativamente homogéneo (aunque algunas bacterias fotosintéticas tienen membranas internas elaboradas). En general, el D.N.A. está enrollado, adherido a la membrana plasmática y concentrado en una región de la célula, llamada nucleoide. Sin embargo, no está físicamente separado del resto del citoplasma por una membrana. Las células procarióticas no tienen organelos como son el núcleo, cloroplastos, ni mitocondrias. Las células bacterianas generalmente son muy pequeñas, con rangos entre los 0.2 y los 10 micrómetros de diámetro.
Las células eucarióticas (“núcleo verdadero”) difieren de las células procarióticas en muchos aspectos. Además de ser más grandes que las células procarióticas (con frecuencia, de más de 10 micrómetros de diámetro), las células eucarióticas contienen una gran variedad de organelos membranosos que le proporcionan a las células una organización estructural y funcional. Técnicamente, el material dentro de la membrana plasmática de una célula eucariótica se divide en el núcleo, un organelo que consta de una membrana de doble capa que contiene al material genético y el citoplasma, que contiene el resto. El citoplasma a su vez está compuesto por varios tipos de organelos, que ocupan casi la mitad del volumen celular, y de una matriz líquida, el citosol (que significa literalmente “solución celular”), el cual contiene los organelos. El citosol, es una solución acuosa de sales, azúcares, aminoácidos, proteínas, ácidos grasos, nucleótidos y otros materiales. Con algunas excepciones, todas las células eucarióticas,
contienen los siguientes organelos, cada uno con sus propias especializaciones
estructurales y funcionales: |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
II.4. Célula vegetal y animal |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Todos los animales y plantas poseen células eucarióticas, pero existen varias diferencias significativas entre las células de los organismos de uno y otro reino. Las células vegetales (véase fig. 12) casi siempre tienen
una pared celular (extra celular) de celulosa. Las células animales
no tienen pared celular. Los hongos y bacterias sí la tienen, pero
no de celulosa. Los plástidos (cloroplastos, leucoplastos y cromoplastos)
son elementos característicos de la mayor parte de las células
vegetales, pero no están presentes en las células animales.
Las células animales (véase fig. 13) invariablemente cuentan con un par de centríolos situados muy cerca del núcleo, por lo regular las células vegetales carecen de ellos.
Las células vegetales, así como las animales, presentan un alto grado de organización, con numerosas estructuras internas delimitadas por membranas; (véase tabla 1). Tanto en las plantas como en los animales, grupos de células similares se organizan en forma de capas o haces laxos llamados tejidos. Estos desempeñan actividades específicas. Los tejidos integran estructuras individuales de forma bien definida conocidas como órganos. Los órganos realizan funciones específicas en el organismo,
(riñones, pulmones, etc.). Así mismo, los órganos
pueden asociarse y construir un aparato orgánico o un sistema complejo,
que lleva a cabo cierta función global.
Tabla núm. 1.Características de los diferentes tipos de células. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
d) Actividad de aprendizaje – Célula vegetal |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
I. Haz una preparación temporal de células del tallo de un bambú de acuerdo con los siguientes pasos: a) Corta con la navaja de un solo filo en la parte plana de la caña. II. Observa el corte de bambú en el lente panorámico. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
e) Actividad de aprendizaje – Célula animal. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 1. Coloca una gota de agua en el centro del portaobjetos. 2. Usando la parte ancha o plana de un palillo de dientes, raspa suavemente sin causarte heridas la parte interna de tu mejilla. 3. Deposita lo obtenido por el palillo sobre la gota de agua y agita suavemente. 4. Añade una gota de lugol o de azul de metileno a la gota de agua que contiene las células epiteliales. 5. Espera un minuto y entonces coloca cuidadosamente el cubreobjetos sobre el material a observar. 6. Inicia la observación con los lentes objetivos de 10X y 40X. 7. Observa atentamente las células del tejido epitelial obtenidas de la parte interna de tu mejilla, pon atención en su forma y estructura. 8. Elabora un dibujo de lo observado con el mayor detalle posible. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
II.5. Pared celular y membrana plasmática |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
“Para permanecer con vida, usted tiene que ser capaz de mantener un equilibrio, un balance, actuar en contra de la entropía, y solo puede llevarse esto a cabo con membranas como las de nuestro mundo” Lewis Thomas, The Lives of a Cell (1974) Por un momento, piense en el reto que representa para un organismo unicelular flotar en un estero, varado por la marea que retrocede. Dicho organismo habita en un medio cuyas condiciones cambian rápida y frecuentemente. Mientras baja la marea, el sol calienta el agua. Conforme el día va pasando, parte del agua se evapora, incrementando la concentración de sales disueltas en la misma. De haber una lluvia repentina o bien al subir la marea, se produce una nueva dilución de sales. Cuando cae la noche, la temperatura del agua desciende. La supervivencia requiere de una barrera protectora entre el interior y el exterior de la célula.Sin embargo, el solo hecho de resistir los cambios ambientales no resulta suficiente. La célula debe absorber luz solar para llevar a cabo la fotosíntesis o ingerir otros organismos a fin de obtener los nutrimentos que necesita para elaborar nuevos componentes celulares. Para mantener un ambiente interno estable en el que sus componentes funcionen mejor, la célula debe mover sales, agua y otras moléculas hacia adentro y afuera. Tiene que excretar productos de desecho, debe detectar claves químicas para la localización de los alimentos, así como condiciones saludables y no saludables. Por ultimo, para reproducirse, debe comunicarse sexualmente por lo menos con otra célula de su propia especie en algún momento de su vida. Una membrana lipídica de sólo dos moléculas de espesor protege el delicado medio interno funcional de la célula y su ambiente externo hostil. La membrana también permite la absorción y liberación selectiva de químicos, en ocasiones mediante el empleo de energía celular. Permite que sucedan reacciones químicas entre una molécula que se encuentra fuera de la célula y otra que se ubica dentro de la misma. Puede deformarse para ingerir alimentos o permitir que la célula se mueva. Todos estos procesos radican en la gran variedad de proteínas que se encuentran embebidas en la membrana lipídica o adosadas a su superficie. Algunos organismos, formados por una célula, como la amiba, sobreviven sin ninguna barrera entre la vida y el medio, solo en su membrana celular. Las células vegetales, las bacterias y muchos protistas, como las diatomeas, tienen paredes por fuera de la membrana celular que les proporcionan sostén mecánico. Las células de los animales multicelulares viven rodeadas por otras células y están inmersas en un líquido parecido a la sangre. Pero todas éstas células poseen membranas celulares muy similares y efectúan varios procesos semejantes por medio de dichas membranas. Paredes celulares La superficie externa de las células de bacterias, vegetales, hongos y algunos protistas está cubierta por una capa dura no viviente, llamada pared celular. Las paredes celulares de los vegetales están compuestas por celulosa y otros polisacáridos, mientras que las paredes celulares de los hongos están compuestas del polisacárido modificado llamado quitina. Las paredes celulares bacterianas tienen una armazón parecido a la quitina a la que están unidas cadenas cortas de aminoácidos y otras moléculas. Las paredes celulares son producidas por la misma célula que rodean. En las plantas, las vesículas membranosas llenas de polisacáridos pegajosos como la pectina (el ingrediente que hace que el jugo de uva forme una jalea) se alinean en el centro de una célula en división. Las vesículas se fusionan, y forman las nuevas membranas plasmáticas que separan a las dos células hijas. Las pectinas previamente formadas y luego contenidas en las vesículas mantienen unidas a las células hijas y dan forma a la lámina media. Las dos células secretan celulosa por sus membranas plasmáticas, por debajo de la lámina media formando la pared celular primaria. Muchas células vegetales posteriormente secretan más celulosa y otros polisacáridos por debajo de la pared primaria para formar una pared celular secundaria gruesa. En algunas células vegetales, la pared secundaria puede hacerse más gruesa que el resto de la célula. Las paredes celulares sostienen y protegen a las células que, de otra forma, serían frágiles. Por ejemplo, las paredes celulares permiten que las plantas y los hongos resistan las fuerzas de la gravedad, el viento y que permanezcan erectas sobre la tierra. Los troncos de los árboles son la prueba fehaciente de la dureza de las paredes celulares, ya que están compuestas casi por completo de celulosa y otros materiales que se han depositado a lo largo de los años y que son capaces de soportar fuerzas impresionantes. Aunque fuertes, las paredes celulares también son porosas, lo que permite el paso fácil de pequeñas moléculas como los minerales, el agua, el oxígeno, el bióxido de carbono, los aminoácidos y azúcares (de otro modo, la célula que se encuentra en su interior pronto moriría). Sin embargo, la estructura que realmente rige las interacciones entre una célula y su ambiente externo es la membrana plasmática.
La membrana plasmática de una célula puede considerarse
como un guardián que solo permite que sustancias específicas
entren o salgan, y que ciertos mensajes pasen del ambiente externo al
interior de la célula. Como guardián, la membrana plasmática
realiza diversas funciones específicas: El modelo del mosaico fluido describe la estructura de la membrana plasmática. De acuerdo con el modelo del mosaico fluido (véase fig. 14) de
la membrana celular desarrollado por los biólogos celulares S.
J. Singer y G. L. Nicolson en 1972, una membrana, vista desde arriba,
parece como un mosaico de azulejos (o tejas) agitados que cambian constantemente.
Una bicapa de fosfolípidos forma el “armazón”
fluido y viscoso para el mosaico, mientras una gran variedad de proteínas
hace el papel de los “azulejos”, que frecuentemente se deslizan
de manera lenta dentro de la bicapa fosfolipídica. Así,
la imagen global cambia lentamente con el tiempo, aunque los componentes
permanezcan relativamente constantes. Aun cuando parezca extraño,
éste modelo refleja la cualidad dinámica de las membranas
reales.
La parte fluida de las membranas es producida por una doble capa de fosfolípidos: la bicapa fosfolipídica. Un fosfolípido consta de dos partes muy diferentes, una cabeza hidrofílica polar y un par de colas hidrofóbicas no polares: todas las células vivas están rodeadas por agua, ya sea de una charca en la que pasa su vida una Amoeba o el líquido extracelular que baña a las células animales. El citoplasma celular casi en su mayor parte está formado por agua. Las membranas plasmáticas, por lo tanto, separan a un citoplasma acuoso de un medio externo acuoso. Bajo tales condiciones, los fosfolípidos se ordenan espontáneamente en una doble capa, llamada bicapa fosfolipídica: Los enlaces de hidrógeno se pueden formar entre el agua y las cabezas de los fosfolípidos, de tal manera que las cabezas ven hacia el citoplasma o hacia el líquido extracelular. Las interacciones hidrofóbicas hacen que las colas fosfolipídicas se escondan dentro de la bicapa. Debido a que las moléculas fosfolipídicas individuales no están unidas entre sí, ésta doble capa es bastante fluida, y las moléculas individuales se mueven con relativa facilidad. En su mayor parte, las moléculas biológicas, como las sales, los aminoácidos y azúcares, son polares y solubles en agua. Por lo tanto, no pueden pasar fácilmente por las colas hidrofóbicas de ácidos grasos, no polares, de la bicapa lipídica. Debido a que la mayor parte de las sustancias que se ponen en contacto con una célula son solubles en agua, la bicapa fosfolipídica actúa como una barrera, evitando la entrada de éstas moléculas. Es en gran parte responsable de la primera de las cuatro funciones de la membrana que mencionamos previamente ---aislar el citoplasma celular del ambiente externo. La bicapa fosfolipídica de las membranas también contiene colesterol. Las membranas de las mitocondrias tienen unas cuantas moléculas de colesterol, pero algunas membranas plasmáticas tienen tanto colesterol como fosfolípidos. El colesterol afecta de diversas formas la estructura de la membrana y su función. Hace que la bicapa sea mas fuerte y flexible, pero menos fluida y permeable a sustancias solubles en el agua como los iones o los monosacáridos. La naturaleza fluida y flexible de la bicapa es muy importante para la función de la membrana. Mientras usted respira, mueve sus ojos o pasa las páginas de este libro, las células de su cuerpo cambian de forma. Si sus membranas plasmáticas fueran rígidas en lugar de flexibles, las células se romperían y morirían. Es más, algunas vesículas de membranas son liberadas constantemente desde el retículo endoplásmico, llegan al aparato de Golgi y nuevamente son liberadas y se fusionan con la membrana plasmática. Las membranas pueden eliminar vesículas y sellarse nuevamente, o fusionarse con vesículas y volver a ser lisas, debido a la naturaleza fluida de la bicapa. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
II.6. Fisiología de la membrana plasmática |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Casi todas las células vivas están bañadas
por un líquido, el cual puede ser el líquido extracelular
del cuerpo humano, el agua de una charca en la cual nada un organismo unicelular,
como la amiba, o las paredes celulares saturadas de agua de una planta joven.
La membrana plasmática separa el citoplasma líquido de la
célula de su ambiente líquido. Iniciaremos nuestro estudio
del transporte de membranas, analizando brevemente las características
de los líquidos, así que empecemos con algunas definiciones.
1. Un fluido es cualquier sustancia que puede moverse o cambiar de forma
en respuesta a las fuerzas externas, sin romperse, se encuentre en estado
líquido o gaseoso. Durante el transporte pasivo, las sustancias se mueven hacia los gradientes de concentración, carga eléctrica o presión. Este movimiento por sí mismo no requiere un gasto de energía. Los gradientes ofrecen la energía potencial para efectuar el movimiento y controlan la dirección del mismo hacia adentro o afuera de la célula. La membrana plasmática actúa como un filtro. Los lípidos y los poros proteicos regulan las moléculas que pueden cruzar e influyen en la frecuencia, así como en la velocidad del movimiento, pero no en la dirección del mismo. En el transporte que requiere energía, las sustancias se mueven en contra de los gradientes de concentración, carga eléctrica o presión. En éste caso, las proteínas de transporte de la membrana plasmática controlan la dirección del movimiento, utilizando energía química a partir del metabolismo celular para originar el movimiento en contra de los gradientes. Una analogía útil para entender la diferencia entre éstos dos tipos de transporte sería considerar lo que sucede cuando se anda en bicicleta. Si no pedalea, sólo puede ir cuesta abajo, como en el transporte pasivo, pero si usted lo hace con suficiente energía también puede ir cuesta arriba. Las moléculas se difunden de las áreas de alta concentración a las de baja concentración. La difusión es el movimiento neto de las moléculas en un
fluido, desde las regiones de alta concentración hasta las de baja
concentración, producida por el gradiente de concentración;
(véase fig. 15). La difusión puede presentarse de una parte
de un fluido a otra, o por una membrana que separa los dos compartimentos
que contienen líquidos. Examinaremos primero el caso más
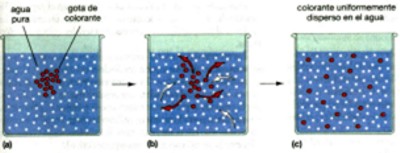
sencillo, la difusión de moléculas dentro de un fluido. Una gota de colorante está compuesta por muchas moléculas
individuales de colorante que se mueven en todas direcciones. Las moléculas
que se desplazan, pero permanecen dentro de la gota de colorante, no cambian
su composición. Pero otras, que se encuentran en el borde de la
gota, debido al movimiento fortuito, escapan hacia el agua. Así,
el movimiento neto de las moléculas de colorante se realiza de
la región de alta concentración de colorante (la gota) hacia
la de baja concentración (el agua). Lo mismo sucede con las moléculas
de agua. El movimiento fortuito ocasiona que algunas se entremezclen con
las moléculas de colorante, y el movimiento neto del agua se da
desde la región de alta concentración de la misma que se
encuentra fuera de la gota hacia la de baja concentración en la
gota.
Para el observador, las moléculas de colorante que se mueven mas allá del borde original de la gota hacen que ésta se vea más grande; las moléculas de agua que invaden la gota diluyen el colorante, provocando que sea más pálida. En primer lugar, cuando la gota es colorante puro y el agua es pura, hay un gradiente de concentración excesivo y el colorante se difunde rápidamente. Conforme disminuyen las diferencias de concentración, el colorante se difunde más y más lentamente. Sin embargo, mientras la concentración del colorante dentro de la gota que se expande es mayor que la concentración del colorante en el resto del vaso, el movimiento neto del colorante será desde la gota hacia el agua, hasta que el colorante se disperse uniformemente en el agua. Entonces, sin ningún gradiente de concentración, ya sea de agua o de colorante, se detiene la difusión. Las moléculas individuales aún se mueven, pero no hay cambios en la concentración ni del agua ni del colorante. Principios de la difusión. 1. La difusión es el movimiento neto de moléculas desde
un gradiente de alta concentración a otro de baja concentración.
Agua, gases disueltos (como el oxígeno, el bióxido de carbono) y moléculas solubles en lípidos (como el alcohol etílico y la vitamina A) se difunden fácilmente al cruzar la bicapa de fosfolípidos. A este proceso se le denomina difusión simple. Por lo general, la rapidez de difusión simple es una función del gradiente de concentración por la membrana, del tamaño de la molécula y de qué tan fácilmente se disuelva en los líquidos (su liposolubilidad): los gradientes de alta concentración, las moléculas de tamaño pequeño y la alta liposolubilidad aumentan la rapidez de difusión simple. Otras moléculas atraviesan la membrana mediante difusión facilitada con ayuda de las proteínas de la membrana. La mayor parte de las moléculas solubles en agua, como los iones (K, Na, Ca), los aminoácidos y los monosacáridos no se pueden mover por la bicapa lipídica. Estas moléculas pueden atravesar y difundirse sólo con la ayuda de dos tipos de proteínas de transporte: las proteínas de canal y las proteínas portadoras. Este proceso recibe el nombre de difusión facilitada. Las membranas plasmáticas contienen relativamente pocos canales
por los que puede pasar cualquier molécula. Por lo tanto, la difusión
facilitada es una función tanto del gradiente de concentración
como de la cantidad de proteínas del transporte. Las moléculas
que cruzan la membrana mediante difusión facilitada generalmente
lo hacen de manera más lenta que las que la cruzan por difusión
simple mediante la bicapa lipídica. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
f) Actividad de aprendizaje – Difusión |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Coloca una pizca de azul de metileno en un vaso de agua
y observa cómo, aunque el líquido esté en reposo, el
colorante se difunde hasta quedar repartido de manera homogénea en
el agua.
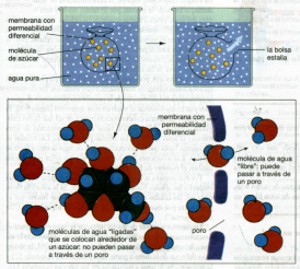
Si te preocupas por medir la cantidad de agua del vaso, su temperatura, el peso del colorante y el tiempo que necesita éste para difundirse uniformemente en el agua, obtendrás una serie de datos útiles como punto de partida para hacer una pequeña investigación. La velocidad de difusión es una variable que acaso esté relacionada con otras variables como la cantidad de agua, la temperatura, la luz. Si te preguntas qué relación puede haber entre la velocidad de difusión y cualquier otra variable, habrás dado el primer paso del método experimental al haber planteado un problema. Ejemplo: ¿Qué relación existe entre la velocidad de difusión y la viscosidad de la fase dispersora? El segundo paso del método, la hipótesis, consistiría en dar una respuesta sobre la posible relación entre esas dos variables. Para comprobar la hipótesis, es necesario elaborar un plan (el diseño experimental) variar la viscosidad de la fase dispersora y ver si altera la velocidad de difusión. Todo ello requiere varios experimentos (realización del trabajo) y gráficas elaboradas con los datos obtenidos. Reunir todas las gráficas en una sola podría ser muy útil para analizar los resultados y obtener una conclusión (pasos quinto y sexto del método experimental). La ósmosis es la difusión de agua por la membrana. El agua, igual que cualquier otra molécula, se mueve mediante difusión de regiones de alta concentración de agua a las de baja concentración. Sin embargo, la difusión del agua que cruza las membranas permeables diferenciales tiene consecuencias tan drásticas que se le ha dado un nombre especial: ósmosis; (véase fig. 16). Analicemos la ósmosis en otro experimento imaginario. Una clase muy simple de membrana con permeabilidad diferencial consta de una hoja impermeable perforada con pequeños poros. Los poros permiten que las moléculas de agua pasen por ellos, pero no moléculas mayores como el azúcar. Suponga que hacemos una bolsa de una membrana como ésta, la llenamos con una solución de azúcar, la amarramos por el extremo y colocamos la bolsa en un vaso de agua pura. La bolsa se hinchará y, si está lo suficientemente débil, estallará. ¿Por qué? Si usted pudiera ver moléculas individuales, notaría que hay dos categorías de moléculas de agua en la solución de azúcar dentro de la bolsa, moléculas de agua “libres”, bien separadas de los azúcares, y moléculas de agua “ligadas”, unidas a los azúcares mediante enlaces de hidrógeno (razón por la que el azúcar se disuelve en el agua). En el agua pura que se encuentra por fuera de la bolsa, claro está que sólo hay moléculas de agua libres. Ahora bien, las moléculas de agua libres se pueden difundir por los poros en la membrana, pero las moléculas de agua ligadas no, porque están unidas, al menos temporalmente, a los voluminosos azúcares. Por lo tanto, la concentración de moléculas de agua libres
es menor dentro de la bolsa que en el agua pura fuera de la misma. Este
gradiente de concentración del agua favorece el movimiento de las
moléculas de agua libres desde el agua pura que se encuentra fuera
de la bolsa hasta la solución de azúcar dentro de la misma.
La bolsa se hincha conforme más moléculas de agua entran
a ella, en comparación con las que la abandonan. El azúcar
de ninguna manera puede escaparse, de tal forma que la concentración
de agua libre dentro de la bolsa siempre es más baja que en el
agua pura fuera de ella. El agua continúa entrando a la bolsa hasta
que revienta.
La presión física que equilibra exactamente la ósmosis
del agua, debido a la diferencia de concentración entre una solución
y el agua pura, se define como la presión osmótica de la
solución. Una solución con presión osmótica
elevada tiene una baja concentración de agua libre; una solución
con una presión osmótica baja tiene una alta concentración
de agua libre. Principios de la ósmosis. 1. La ósmosis es la difusión de agua por una membrana con
permeabilidad diferencial.
En su mayor parte las membranas plasmáticas son muy permeables al agua. Debido a que todas las células contienen sales disueltas, proteínas, azúcares, etc., el flujo de agua por la membrana plasmática depende de la concentración de agua en el líquido que baña las células. Los fluidos extracelulares de los animales generalmente son isotónicos (“de la misma fuerza”) hacia el interior de las células corporales; esto es, la concentración de agua es igual dentro o fuera de la misma. Si los eritrocitos, por ejemplo, se sacan del cuerpo y se sumergen en
soluciones salinas de concentraciones variables, los efectos de la permeabilidad
diferencial de la membrana plasmática al agua y a las partículas
disueltas se vuelven muy aparentes. Si la solución tiene una concentración
mayor de sal que la del citoplasma (esto es, si la solución tiene
una concentración de agua menor) el agua saldrá de las células
por ósmosis. Las células se encogerán hasta que las
concentraciones de agua dentro y fuera se igualen. Las soluciones que
hacen que el agua salga por ósmosis recibe el nombre de hipertónicas
(“de mayor fuerza”). La ósmosis a través de las membranas es importante para
el funcionamiento de muchos sistemas biológicos, incluyendo la
ingesta de agua por las raíces de las plantas, la absorción
del agua de la dieta a partir de los intestinos y la reabsorción
de agua y minerales en los riñones. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
g) Actividad de aprendizaje – Ósmosis. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Coloca una hoja de Anacharis (comúnmente conocida
como elodea) y colócala entre un portaobjetos y un cubreobjetos.
Si añades agua de la llave, la hoja estará en un medio isotónico
(de igual concentración) favorable para hacer las observaciones.
Lo que aparecerá en el campo, si has enfocado correctamente la preparación,
será una serie de células de forma rectangular provistas en
su interior de numerosos granos verdes: los cloroplastos.
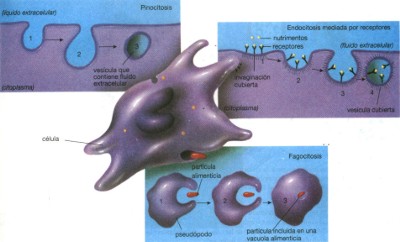
La estructura que enmarca a la célula es la cápsula de secreción o pared celular, que por ningún motivo debe confundirse con la membrana fundamental. A diferencia de la de la membrana celular, la cápsula de secreción resulta perfectamente visible con el microscopio ordinario, es rígida y está formada principalmente por celulosa. Tan rígida es esta cápsula que gracias a ella pueden las plantas mantenerse erguidas, de ahí la tendencia a considerar a la cápsula de secreción como el esqueleto de los vegetales. ¿Quiere esto decir que las células animales carecen de cápsula de secreción? Efectivamente, aunque justo es señalar la existencia de células excepcionales capaces de producir una estructura llamada cubierta externa, como ocurre con el óvulo. Pero es menester recordar que, sin excepción, las células (vegetales, animales, protistas o moneras) tienen membrana celular. Si en un extremo del portaobjetos colocas un pedazo de papel filtro para extraer el agua de la preparación y por el lado opuesto añades agua destilada, las moléculas de agua empezarán a entrar por ósmosis en las células, las cuales reventarían si no fuera por la cápsula de secreción. En tal caso se dice que la célula está en solución hipotónica; este fenómeno se conoce como turgencia. En cambio, si sustituyes el agua destilada por una solución saturada de sal, las células pierden agua al grado de que la membrana fundamental queda separada de la cápsula de secreción. Entonces la célula se encuentra en un medio hipertónico; este proceso biológico recibe el nombre de plasmólisis. La célula de Anacharis (elodea) es un espécimen adecuado para desarrollar ambas observaciones. El transporte activo utiliza energía para mover moléculas y cruzar la membrana. En el transporte activo, las proteínas de membrana utilizan la energía celular para mover moléculas individuales y cruzar la membrana plasmática, generalmente en contra de su gradiente de concentración. Las proteínas de transporte activo con frecuencia reciben el nombre de bombas, en una analogía con las bombas de agua porque utilizan energía para mover moléculas “hacia arriba” en contra de un gradiente de concentración. Las bombas de la membrana plasmática son vitales en los vegetales para absorber minerales, en la absorción de minerales en los intestinos y para mantener los gradientes de concentración esenciales para el funcionamiento de una neurona. Las células engloban partículas con líquidos mediante la endocitosis. Las células pueden obtener líquidos o partículas, especialmente proteínas grandes o microorganismos completos como las bacterias, mediante un proceso, llamado endocitosis (del griego “dentro de la célula”). Durante la endocitosis, la membrana plasmática engloba la partícula o la gota de líquido y emite un saco membranoso, denominado vesícula, con la partícula o líquido adentro y la lleva al interior del citoplasma. Pueden distinguirse tres tipos de endocitosis, basados en el tamaño de la partícula obtenida y el método de obtención; (véase fig. 17). La pinocitosis introduce líquido dentro de la célula. En la pinocitosis (“la forma de beber de la célula”) o endocitosis de fase líquida, una parte muy pequeña de membrana plasmática se hunde conteniendo fluido extracelular, y lo introduce en el citoplasma como una pequeña vesícula. La pinocitosis mueve una gota de fluido extracelular, contenida dentro de la parte que se hunde dentro de la membrana, hacia el interior de la célula. Por lo tanto, la célula obtiene materiales en proporción a su concentración en el líquido extracelular. La endocitosis mediada por receptor, mueve moléculas específicas al interior de la célula. La célula puede captar ciertas moléculas (por ejemplo colesterol) más eficientemente por medio de un proceso conocido como endocitosis mediada por receptor. La mayor parte de las membranas plasmáticas cuenta con muchos receptores proteicos en sus superficies externas. En casi todos los casos, éstos receptores se mueven por la bicapa fosfolipídica y se acumulan en depresiones de la membrana plasmática llamadas fosas cubiertas. Si la molécula correcta se pone en contacto con un receptor proteico en una de ésas fosas cubiertas, se fija al sitio de unión. La fosa cubierta se profundiza en una bolsa en forma de U que finalmente queda dentro del citoplasma como una vesícula cubierta. Tanto el complejo de nutrimento—receptor como un poco de fluido extracelular quedan dentro de la célula en la vesícula cubierta.
La fagocitosis (“la forma de comer de la célula”)
se utiliza para captar partículas grandes, incluidos microorganismos
completos. Por ejemplo, cuando una Amoeba siente apetitoso a un Paramecium,
emite extensiones de su membrana superficial, llamadas pseudópodos
(del latín “falso pié”). Los pseudópodos
rodean al infortunado Paramecium, sus extremos se fusionan y la presa
es llevada al interior de la Amoeba para su digestión. La vesícula
resultante, llamada vacuola alimenticia, se fusiona con lisosomas cuyas
enzimas digieren a la presa. Los leucocitos también utilizan la
fagocitosis y la digestión intracelular para englobar y destruir
bacterias que han invadido su organismo.
La exocitosis expulsa material de la célula. Lo contrario de la endocitosis es la exocitosis (del griego “fuera
de la célula”), que con frecuencia es utilizada por parte
de las células para deshacerse de materiales no deseados, productos
de desecho de la digestión o para secretar materiales, que pueden
ser hormonas, hacia el fluido extracelular. Durante la exocitosis, una
vesícula creada por el aparato de Golgi se mueve a la superficie
celular, en la que la membrana de la vesícula se fusiona con la
membrana plasmática. La vesícula se abre al fluido extracelular
y su contenido se difunde hacia fuera. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
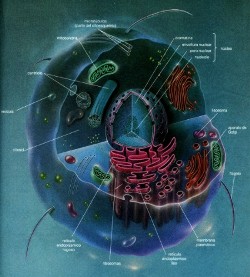
II.7. Citoplasma y núcleo. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Citoplasma.
Al contenido interno de la célula a veces se le denomina protoplasma. Algunos autores han considerado que es la sustancia de la vida, pero como ya se dijo, la célula es la unidad de la vida, y una parte constituyente no está viva por sí misma. Otra desventaja de utilizar el término protoplasma, es que sugiere una uniformidad de composición, nada mas lejos de la verdad. Nuestro protoplasma difiere del protoplasma de las células del perro, el caballo, el gato y la amiba; es diferente de los plasmas de todos los demás organismos. El protoplasma varía en composición y propiedades, no solo entre, sino dentro de los organismos; el protoplasma de los eritrocitos difiere del de una célula ósea, cerebral o hepática. Se utiliza el término protoplasma, porque es una palabra descriptiva, adecuada para indicar el material que se haya en la célula; sin embargo, no debe considerársele como una entidad químicamente definida o caracterizada por una vida propia, porque esto no tiene sentido. El protoplasma de la célula que está rodeado por la membrana
plasmática, pero que está fuera del núcleo de la
célula, se llama citoplasma. Químicamente, consta de proteínas
y lípidos, algunos carbohidratos, minerales, sales y bastante agua
(de 70 a 90%). La proporción de éstos componentes varía
mucho de una célula a otra y de organismo a organismo. El citoplasma
es un líquido viscoso coloidal. ¿Qué es un coloide?.
Una sustancia coloidal familiar es la gelatina, hecha de los huesos, tendones
y ligamentos del ganado bovino mediante un tratamiento con agua caliente
para hacer soluble algunos de sus componentes. Es una proteína,
y cuando se le pone en agua caliente, las moléculas de proteína
se dispersan fina y uniformemente en el agua y así permanecen debido
a su carga electrostática y a que son bombardeadas por la constante
actividad de las moléculas de agua. La gelatina tiene otra propiedad
familiar al químico casero: puede existir como un sólido
o como un líquido. Si se coloca un poco de gelatina caliente disuelta
en un lugar frío, se solidifica. El cambio del estado líquido
al sólido se llama transformación sol-gel (solución
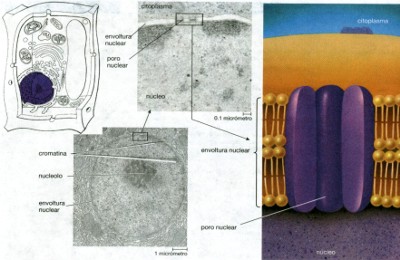
–sólido). El material coloidal del citoplasma de las células está compuesto principalmente de moléculas de proteína y glóbulos de grasa. El citoplasma también contiene materiales más grandes que las partículas coloidales, y tales partículas no permanecen estables. Si se dejaran a la fuerza de la gravedad, éstas partículas se asentarían en el tiempo. A los sistemas que contienen materiales semejantes se les denominan suspensiones. Hay suspensiones de cristales y de otros materiales en forma de partículas que se encuentran dispersas ordinariamente en las soluciones coloidales acuosas del citoplasma. Dentro de la célula existe material en solución; esto es, disuelto mas o menos uniformemente en el agua de la célula. El tamaño de tales partículas puede tener el tamaño de átomos, moléculas o de pequeños grupos de éstos. Las moléculas de éste tamaño no pueden verse con el microscopio, no se depositan por gravedad, no pueden separarse mediante la filtración y permanecen dispersas por la constante agitación de las moléculas de agua. Además, se hallan disueltas en el citoplasma celular sustancias gaseosas como dióxido de carbono y oxígeno; éstos son con frecuencia los componentes menos abundantes del citoplasma. En el citoplasma están suspendidos los organelos y las micro estructuras celulares. Núcleo. Descubierto por Robert Brown en 1831. Es el centro de control de la célula. El ácido desoxirribonucleico (DNA) es el material genético de las células vivas. El DNA de una célula es la información necesaria para construir a la célula y dirigir las incontables reacciones químicas necesarias para la vida y la reproducción. De igual manera que un plano se utiliza selectivamente para construir cada parte de una casa, la información genética en el DNA es empleada de manera selectiva por la célula, dependiendo de su estado de desarrollo y sus condiciones ambientales. En las células eucarióticas, el DNA está dentro del núcleo; (véase fig. 18). El núcleo consta de tres componentes fácilmente distinguibles.
La envoltura nuclear separa el material nuclear del citoplasma. Dentro
de la envoltura nuclear, el núcleo contiene un material granular
llamado cromatina, y una región más obscura llamada nucleolo.
El núcleo está aislado del resto de la célula por una envoltura nuclear formada por dos membranas interrumpidas por poros. Los poros nucleares son estructuras complejas que contienen por lo menos ocho subunidades proteicas con un canal pequeño en el centro. El agua, los iones y las moléculas pequeñas como el ATP pueden pasar libremente por el canal central, pero éste regula el paso de moléculas mayores, en especial de proteínas y de RNA, probablemente conforme se mueven las unidades proteicas ya sea para agrandar o restringir el acceso al canal. De éste modo los poros ayudan a controlar el flujo de información de y desde el DNA.
En una micrografía electrónica el núcleo muestra una apariencia granular, con regiones más obscuras y más claras pero sin una estructura obvia. Debido a que el núcleo se colorea intensamente por varios colorantes comunes, utilizados en la microscopía de luz, los primeros microscopistas llamaron al material nuclear cromatina (que significa “substancia coloreada”). Los biólogos han aprendido que la cromatina está compuesta de DNA y proteínas. Aunque no podemos verlos en las micrografías electrónicas ordinarias, el DNA eucariótico y las proteínas forman hebras largas, llamadas cromosomas (“cuerpos coloreados”). Cuando las células se dividen, cada cromosoma se enrolla, haciéndose más grueso y más corto. Los cromosomas “condensados” que resultan son más fáciles de ver con microscopios de luz. Los procesos celulares (las reacciones químicas responsables del crecimiento, la reparación, la nutrición, la captación de energía y su uso, y la reproducción) están guiadas por la información codificada en el DNA. Debido a que el DNA permanece en el núcleo, ya que la mayor parte de las reacciones químicas que controla suceden en el citoplasma, deben intercambiarse moléculas entre el núcleo y el citoplasma. La información genética que contiene el DNA se copia en moléculas de RNA, éstas pasan por los poros de la cubierta nuclear hacia el citoplasma. Esta información se utiliza para dirigir la síntesis proteica. Estas proteínas incluyen enzimas que regulan las reacciones químicas, proteínas de la membrana que coordinan las interacciones entre la célula y su medio, y la gran variedad de proteínas estructurales. Algunas de éstas proteínas pasan del citoplasma al núcleo y regulan la transferencia de información del DNA al RNA, dependiendo de lo que esté sucediendo en el citoplasma y en el medio extracelular.
La mayor parte de los núcleos eucarióticos tienen una
o más regiones que se tiñen de color más obscuro,
llamados nucleolo (en la figura 18 a se muestra un nucleolo, “pequeño
núcleo”), el cual es uno de los sitios de la síntesis
de los ribosomas. Un ribosoma es una pequeña partícula compuesta
de RNA y proteínas que sirve como “mesa de trabajo”
para sintetizar proteínas. Al igual que muchas mesas de trabajo,
los ribosomas no son específicos --- el mismo conjunto de herramientas
puede utilizarse para construir muchos objetos diferentes. Cualquier ribosoma
puede utilizarse para sintetizar cualquiera de los cientos de proteínas
que se producen en una célula. En las micrografías electrónicas,
los ribosomas aparecen como gránulos obscuros, ya distribuidos
en el citosol o adosados a la membrana de la capa nuclear y del retículo
endoplásmico. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
h) Actividad de aprendizaje – Núcleo y nucleolo |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Las células de cebolla se usan para la observación
del núcleo y el nucléolo. Estos dos organelos celulares se
encuentran en células vivas. Puede haber varios nucléolos
que aparecen como pequeños puntos dentro del núcleo celular.
El núcleo es un organelo celular redondeado dentro de la célula.
Trabaja de acuerdo con los siguientes pasos para hacer una preparación temporal de cebolla. I. Quita la mitad de una hoja de cebolla. Coloca un cubreobjetos sobre la preparación, y golpéalo suavemente con el borrador de tu lápiz o pluma para eliminar las burbujas de aire. V. Observa las células con el objetivo panorámico y con
el seco fuerte. La preparación tiene el aspecto de una pared de
ladrillos, donde los ladrillos son las células y la pared celular
es la que los une o los separa. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
II.8. Organelos citoplasmáticos |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
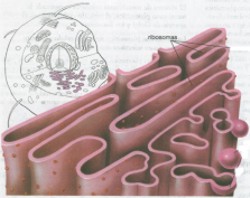
| Retículo endoplásmico y Ribosomas. En
1945 Keith Porter descubrió un sistema de membranas internas en
el citoplasma al que denominó retículo endoplásmico;
(véase fig. 19).
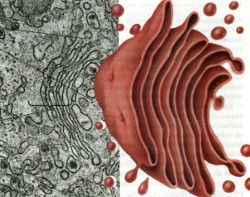
Los ribosomas que se encuentran sobre el RE también fabrican las proteínas que algunas células secretoras exportan hacia sus alrededores, incluyendo las enzimas digestivas y las hormonas proteicas (por ejemplo, insulina secretada por las células del páncreas). Conforme los ribosomas que se encuentran en la cara externa del RE, sintetizan estas proteínas, son transportadas de inmediato dentro de los canales. Las proteínas se mueven por el RE y se acumulan en los extremos, especialmente los extremos cercanos al aparato de Golgi. Entonces estos cúmulos son “eliminados” y forman vesículas recubiertas por membrana que migran hacia el aparato de Golgi. Aparato de Golgi. En 1898, el científico italiano Camilo Golgi observó una
serie de vesículas en las células nerviosas de una lechuza
bodeguera. Estas vesículas recibieron el nombre de aparato de Golgi
en honor a su descubridor; (véase fig. 20).
El aparato de Golgi es un conjunto especializado de sacos membranosos que se originan a partir del retículo endoplásmico. De hecho, el aparato de Golgi se parece a una pila de RE liso que ha sido usada, aplanando la parte central y haciendo que los extremos sobresalgan. Las vesículas eliminadas se fusionan con los sacos en uno de los lados de la pila del Golgi, agregando su membrana a la del Golgi y vaciando su contenido en los sacos del aparato de Golgi. En el lado opuesto de la pila son eliminadas del Golgi otras vesículas, que contienen proteínas, lípidos y otras moléculas complejas. El aparato de Golgi realiza las siguientes tres funciones principales: 1. Separa las proteínas y los lípidos que se reciben del
RE de acuerdo con su destino; por ejemplo, el Golgi separa las enzimas
digestivas que son limitadas por los lisosomas a partir de las hormonas
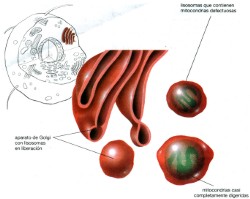
que serán secretadas por la célula. Lisosomas. Funcionan como el sistema digestivo de la célula.
Los lisosomas reconocen estas vacuolas alimenticias y se fusionan con ellas. El contenido de las dos vesículas se mezcla y las enzimas lisosómicas digieren los alimentos convirtiéndolos en aminoácidos, monosacáridos, ácidos grasos y otras moléculas pequeñas. Estas moléculas simples luego se difunden fuera del lisosoma hacia el citosol para nutrir la célula. Los biólogos celulares continúan buscando el mecanismo por el cual los lisosomas reconocen estas vacuolas alimenticias. Los lisosomas también digieren organelos defectuosos o que no
funcionan de manera adecuada, como las mitocondrias o los cloroplastos.
Después de identificar estos organelos, la célula los incluye
en vesículas elaboradas de membrana en el RE. Estas vesículas
se fusionan con los lisosomas, y las enzimas digestivas dentro de estos
últimos permiten que la célula recicle materiales valiosos
a partir de organelos difuntos. Cómo identifica la célula
a los organelos que ya no son útiles es aún un tema de investigación. Los peroxisomas se parecen a los lisosomas, excepto que las enzimas de los peroxisomas tienen funciones oxidativas. Estos últimos intervienen en la desaminación oxidativa de los aminoácidos, reacción indispensable para la conversión de proteínas en otros tipos de compuestos. Cloroplastos y mitocondrias: Cada célula tiene una gran necesidad de energía para fabricar materiales, tomar cosas del medio y eliminar otras, así como para moverse y reproducirse. Como señaló Lewis Thomas, “hay estructuras que se tuercen dentro de cada una de nuestras células y proporcionan toda la energía para vivir”. Éstas son los cloroplastos y las mitocondrias, “criaturas esencialmente extrañas”, de las cuales se piensa que tuvieron que haber evolucionado de bacterias que se instalaron hace mucho tiempo dentro de una célula eucariótica. Los cloroplastos y las mitocondrias son parecidos en muchos aspectos. Ambos generalmente son oblongos, de 1 a 5 micrómetros de longitud aproximadamente y están rodeados por una membrana doble. Ambos tienen enzimas que sintetizan ATP, aunque los sistemas sean utilizados de muy diferentes maneras. Finalmente, los dos tienen muchas características, incluido su propio DNA, remanente de su probable evolución de los organismos de vida libre. Sin embargo, también hay muchas diferencias que corresponden a la gran cantidad de papeles distintos en las células: los cloroplastos captan la energía de la luz solar durante la fotosíntesis y la almacenan en un azúcar, mientras que las mitocondrias convierten la energía de la luz en ATP para su uso en la célula. Los cloroplastos son el sitio en el que se efectúa la fotosíntesis. Los cloroplastos solo se encuentran en los vegetales y en algunos protistas,
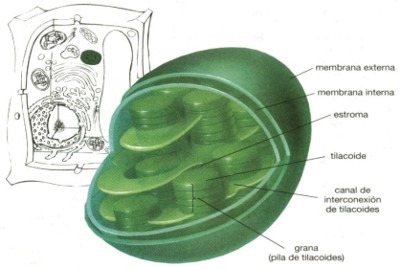
en especial las algas unicelulares; (véase fig. 22). Los cloroplastos
están rodeados por dos membranas, aunque hay muy poco espacio entre
ellas. La membrana interna contiene un material semifluido, llamado estroma.
Contenidas dentro del estroma, están las pilas interconectadas
de sacos membranosos vacíos. Los sacos individuales reciben el
nombre de tilacoides, y a una pila de sacos se le denomina grana. Durante
la formación de los cloroplastos, los tilacoides probablemente
se desprendieron de la membrana interna dentro del estroma; en un cloroplasto
maduro, los tilacoides están conectados a la membrana interna.
Las membranas tilacoides contienen un pigmento verde, la clorofila (que
le da a la planta su color verde ), así como otras moléculas
pigmentadas. Durante la fotosíntesis, la clorofila capta la energía
de la luz solar y la transfiere a otras moléculas de las membranas
tilacoides. Estas moléculas a su vez transfieren la energía
al ATP y a otras moléculas portadoras de energía. Las portadoras
de energía se difunden dentro del estroma, en el que |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
i) Actividad de aprendizaje - Cloroplastos |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Otros organelos celulares presentes en las células
vegetales son los cloroplastos. En la Elodea, que es una planta acuática,
se pueden observar estas importantes estructuras.
I. Haz una preparación temporal de una hoja de Elodea.
Cromoplastos. Plastos que dan a muchas flores y hojas sus colores amarillo,
naranja o rojo; contienen pigmentos. Leucoplastos. Sirven como depósito de alimentos para la célula y contienen aceite, granos de almidón y proteínas. Por ejemplo, las papas son masas de células llenas de plástidos que contienen almidón. Las vacuolas realizan muchas funciones, incluidas las de soporte, almacenamiento
y eliminación de alimentos y desechos. La célula vegetal almacena sustancias en los líquidos de la vacuola, entre ellas aminoácidos, proteínas, sales, carbohidratos y el pigmento rojo antocianina (el rojo de las rosas, cebollas moradas y betabeles). Las mitocondrias producen ATP mediante la utilización de la energía
almacenada en las moléculas alimenticias.
Microfilamentos. Los microfilamentos permiten que las células cambien de forma y guíen el movimiento de los organelos. Los microfilamentos son haces que consisten principalmente de la proteína actina, en ocasiones asociada con una segunda proteína, la miosina. La miosina tiene pequeñas extensiones que se pueden asir a la actina y la flexionan, deslizando el filamento de actina sobre el de miosina, igual que un marinero que jala un ancla con una cuerda, y pone una mano tras la otra. La función más conocida de la actina y la miosina es la contracción muscular, pero los microfilamentos de actina y miosina también contribuyen a los cambios de forma y al movimiento de la mayor parte, si no es que de todos, los organelos de las células eucarióticas.
Hay por lo menos cinco tipos de filamentos intermedios, cada uno compuesto por una proteína diferente. Cada variedad de filamento intermedio por lo general se encuentra en un solo tipo o en algunos tipos celulares. Los filamentos intermedios realizan una gran variedad de funciones, en general son armazones que proporcionan forma a las células y fijan varias de sus partes, por ejemplo, el axón de una neurona está sostenido por un esqueleto interno de filamentos intermedios asociados con microtúbulos. En muchas células, al menos algunas de las proteínas de la membrana plasmática, están adheridas a filamentos intermedios. Por último, los filamentos intermedios sujetan a los microfilamentos de actina en las células musculares, garantizando que las células no se separen durante las contracciones intensas.
Los microtúbulos ayudan a la posición, la fijación y movimiento de organelos, y alteran las formas celulares. Los microtúbulos son cilindros huecos formados por muchas subunidades en forma de mancuerna de gimnasia, de la proteína tubulina. Algunos microtúbulos son permanentes, como los que conforman los cilios y los flagelos, estructuras que utilizan muchas células para moverse. Sin embargo, varios microtúbulos son transitorios y se ensamblan y se desensamblan conforme la célula lo requiere. Todas las células eucarióticas tienen un centro organizador de microtúbulos cerca del núcleo. Durante la división celular, éste centro produce una estructura en forma de balón de fútbol americano, formada por microtúbulos, que recibe el nombre de huso mitótico y moviliza los cromosomas hacia las dos células hijas. En las células animales, se encuentra un par prominente de centríolos;
(véase fig. 24) en el centro organizador de microtúbulos
cercano al núcleo. Los centríolos son anillos cortos en
forma de barril, formados por microtúbulos. Cuando una célula
se divide, los centríolos se duplican y solo un par se mueve hacia
cada célula hija.
Con frecuencia, los microtúbulos se asocian con pequeñas proteínas que viajan a lo largo de ellos, como las locomotoras en una vía férrea. Estas proteínas se unen a diversos organelos y los llevan hacia el lugar adecuado en la célula, donde lo fijan utilizando al microtúbulo como un andamio. El retículo endoplásmico y el aparato de Golgi están colocados en su sitio mediante microtúbulos. Los movimientos de los organelos y de los cromosomas durante la división celular también son producidos por las proteínas que se mueven a lo largo de los microtúbulos. El ensamblado y el desensamblado de los microtúbulos contribuyen a los cambios en las formas generales de las células, por ejemplo, cuando las células nerviosas de la médula espinal se prolongan hacia los músculos de las piernas.
Los cilios y flagelos mueven a la célula o a los fluidos que pasan
por ella.
Pequeños “brazos” proteicos se proyectan desde cada par de microtúbulos en el anillo externo. Estos brazos se unen al par vecino de microtúbulos y lo flexionan, ocasionando el movimiento del primer par en relación con el segundo. Sin embargo, el cuerpo basal fija firmemente la “base” de todos los microtúbulos en el cilio o el flagelo. La energía del ATP potencia el movimiento de los brazos proteicos durante el movimiento de los microtúbulos. Los cilios y flagelos se mueven casi continuamente y necesitan grandes cantidades de ATP, las cuales se generan por las mitocondrias que generalmente se encuentran en abundancia cerca de los cuerpos basales. Las principales diferencias entre los cilios y los flagelos radican
en cuanto al número, la longitud y la dirección de la fuerza
que generan. En general, los cilios (del latín “pestaña”)
son cortos (de 10 a 25 micrómetros de longitud) y numerosos. Proporcionan
fuerza en una dirección paralela a la membrana plasmática,
como los remos en una canoa. Esto se efectúa por medio de un movimiento
“en filas” bastante rígido durante el llamado golpe
efectivo, presentándose la mayor inclinación en la base
del cilio, y un golpe de retorno flexible. Los flagelos (del latín
“látigo”) son largos (de 50 a 75 micrómetros),
generalmente poco numerosos y proporcionan una fuerza perpendicular a
la membrana plasmática, como el motor en una lancha. Algunos organismos unicelulares, como el Paramecium y la Euglena, utilizan los cilios y los flagelos para movilizarse. La mayor parte de los espermatozoides animales también se mueven mediante flagelos. En animales multicelulares, los cilios ocasionalmente se utilizan para la locomoción, moviendo al animal dentro de un líquido. Muchos invertebrados acuáticos pequeños, por ejemplo, nadan por medio del movimiento coordinado de filas de cilios, como los remos de un galeón romano. Las células ciliadas se encuentran en límites de estructuras
tan diversas como las branquias de los ostiones (para el movimiento de
agua rica en alimentos y oxígeno), los oviductos de los mamíferos
hembra (para el movimiento de los óvulos por el oviducto y hacia
el útero), y los tractos respiratorios de casi todos los vertebrados
terrestres (limpiando restos y microorganismos de las vías aéreas
y los pulmones). |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
II.9. Ciclo celular eucariótico – mitosis |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Es una paradoja de la naturaleza que los seres vivos cambien
y, sin embargo, permanezcan los mismos. Considerada toda la vida, podemos
reconocer que envejecemos y morimos; pero en una escala de tiempo más
corta, digamos minutos u horas, difícilmente notamos alguna diferencia
en nuestro aspecto: somos los mismos. O ¿en realidad lo somos? Un
examen crítico nos muestra que no somos los mismos que hace un instante.
Nuestros cuerpos están en constante actividad, y el cambio es interminable.
Las células de las que estamos compuestos trabajan continuamente:
se reproducen, crecen, se especializan y mueren. Nuestras células
deben de trabajar mucho para conservar el status quo, porque deben reemplazar
a las que se pierden por el uso. Ninguna célula vive eternamente,
y cada segundo mueren unos 50 millones de células del cuerpo; durante
éste periodo, se produce la misma cantidad de células nuevas
para tomar su lugar. Las células, como a las personas, varían
en el tiempo de su ciclo de vida; una célula intestinal vive 36 horas,
luego muere y sale del cuerpo; un leucocito vive unas dos semanas; un eritrocito,
cerca de cuatro meses, y una célula nerviosa, 60 años o más.
¿Qué tipo de delicado equilibrio opera en nuestro cuerpo para preservar nuestra integridad? ¿Cómo es posible que las células activas del cuerpo, que pueden sumar millones de millones, puedan regularse? Y ¿cómo es que a veces ciertas células pierdan su control reglamentario y no se sometan ya a los patrones de regulación ordinarios, convirtiéndose en células cancerosas, sin ley? Comenzamos la vida como una sola célula y, a través de acontecimientos notorios, esa célula se dividió en muchas células más pequeñas. Más tarde, utilizando materias primas suministradas desde el exterior, aumentaron de tamaño, se multiplicaron de nuevo, y pronto fueron millares; conforme proseguía el proceso, se propagaron y finalmente fueron millones de células: se organizaron en nerviosas, epiteliales, hepáticas y otras más distintas entre sí. ¿Cómo reconocieron las células originales que su función era la multiplicación? ¿Cuándo recibieron la señal de cesar de dividirse continuamente y equilibrarse con la muerte celular? ¿Por qué, y cómo se especializaron nuestras células? Antes de poder responder preguntas como éstas, debemos comprender una actividad básica de la célula viva: Su reproducción. Las células se reproducen dividiendose. El ciclo celular eucariótico consta de dos fases principales.
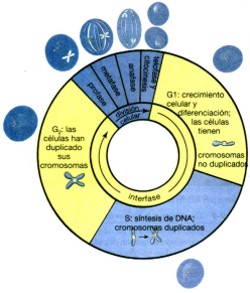
La mayor parte de la vida de una célula (el 90 por ciento o más en general) la pasa en interfase. Los biólogos celulares dividieron a la interfase en tres subfases, llamadas G1 (el “primer nexo” con la síntesis de DNA), G1 (cuando se efectúa la síntesis de DNA) y G2 (el “segundo nexo” con la síntesis de DNA; véase fig. 26). El periodo posterior a la división celular más reciente y previa a la duplicación de los cromosomas es la fase G1. Aunque la designación de “primer nexo” en la síntesis de DNA hace suponer que la célula está descansando antes de proceder a sintetizar el DNA, no sucede así. De hecho, la mayor parte del crecimiento y de la actividad de la célula ocurre durante la fase G1. La célula adquiere nutrimentos de su medio, lleva a cabo sus funciones especializadas (por ejemplo, síntesis y secreción de hormonas) y crece. Más tarde, durante la fase G1, en un punto llamado de restricción, en la célula ocurre una especie de “evaluación interna” de su capacidad para completar el ciclo celular y producir dos células hijas viables. Si la evaluación resulta negativa, la célula no se divide; si resulta positiva, la célula está autorizada para duplicar el DNA y entrar en la división celular. La duplicación de cromosomas defina la S, o fase de síntesis, ya que éste es el único momento de síntesis de DNA que ocurre en condiciones normales. Cada cromosoma se duplica—sólo una vez. Durante la fase S, las células animales también duplican sus centríolos. El periodo posterior a la síntesis de DNA, pero previo a la próxima división celular, es la fase G2. La célula ya está autorizada para efectuar la división celular antes de hacerlo en la fase G2. Durante la mayor parte de la G2 se sintetizan las moléculas necesarias de DNA para la división celular. Muchos tipos de células de mamíferos progresan lentamente durante la interfase, pasando cinco horas en la etapa G1, aproximadamente, siete duplicando su DNA durante la fase S, y tres en G2 preparándose para la división. Aunque la división celular generalmente dura en promedio una hora, algunos tipos celulares tienen ciclos celulares muy cortos, mientras que otros pueden durar semanas o toda su vida sin dividirse. Las divisiones celulares tempranas de un embrión animal ocurren en una sucesión rápida casi carente por completo de la fase G1 y, por lo tanto, casi no hay crecimiento entre ambas divisiones. Por el contrario, las neuronas del cerebro de los mamíferos adultos no se dividen y permanecen en la fase G1 durante toda su vida. La división celular consiste de una división nuclear y de una división citoplasmática. Durante la división nuclear, copias completas e idénticas
de todos los cromosomas quedan contenidas en dos núcleos nuevos.
La división nuclear recibe el nombre de mitosis (del griego “hilo”)
porque los cromosomas tienen estructuras parecidas a hilos en ése
momento.
Durante la división citoplásmica, la citocinesis, el citoplasma se parte en dos células hijas, recibiendo cada célula uno de los núcleos recientemente formados y, en general, cantidades casi iguales de citoplasma. Aunque parezca increíble, una célula nerviosa y un hepatocito
tienen los mismos genes. Sus diferencias en estructura y función
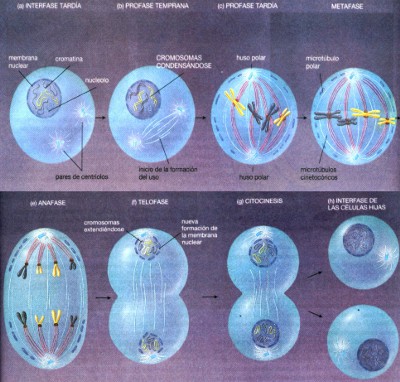
se deben al diferente uso de los genes durante la vida de la célula. Como señalamos, la división celular eucariótica consta de mitosis (división nuclear) y citocinesis (división citoplasmática). En general éstos dos procesos suceden juntos pero, en algunas células, la mitosis puede presentarse sin citocinesis. Antes que iniciemos nuestro análisis de la mitosis, recuerde que los cromosomas se duplican durante la fase S de la interfase. Por lo tanto, cuando se inicia la mitosis, cada cromosoma ya consta de dos cromátidas hermanas unidas por el centrómero. Por conveniencia, la mitosis se divide en cuatro fases: profase, metafase, anafase y telofase (véase figuras 27 y 28). Sin embargo, como suceden en casi todos los procesos biológicos éstas fases no son realmente acontecimientos independientes, por el contrario, forman un suceso continuo, en el que cada fase da lugar a la siguiente. Durante la profase los cromosomas se condensan, se forma el huso mitótico y se fija a los cromosomas. La primera fase de la mitosis recibe el nombre de profase (del griego
“etapa previa”). La profase se presenta después de
la fase G2 de la interfase y suceden tres hechos principales: la condensación
de los cromosomas, el ensamble del huso mitótico y la captura de
los cromosomas por parte del huso.
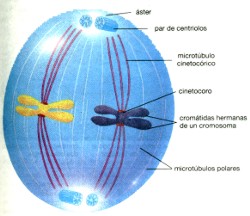
Durante la profase temprana, los cromosomas se condensan y el nucleolo desaparece. Durante la profase, los cromosomas se condensan y se vuelven visibles a la microscopía óptica. Conforme los cromosomas se condensan, los nucleolos desaparecen. El nucleolo contiene un conglomerado, formado por partes de varios cromosomas que participan en la síntesis de ribosomas. Conforme se condensan y separan los cromosomas, se rompe éste conglomerado. Hacia el final de la profase, ya que los cromosomas se han condensado, se empieza a formar el huso mitótico que es un arreglo elipsoide que consta de microtúbulos y se origina de la siguiente manera. Durante la interfase, una célula contiene un arreglo de microtúbulos que parten del centro organizador de microtúbulos que se encuentra cerca del núcleo. Durante la profase, los microtúbulos de la interfase se desintegran en subunidades individuales de la proteína tubulina. El centro organizador de microtúbulos se parte en dos y cada centríolo hijo empieza a formar un nuevo arreglo de microtúbulos a partir de las subunidades de tubulina. Este nuevo arreglo de microtúbulos recibe el nombre de huso mitótico debido a la forma ahusada que presenta—gruesa en la parte central y delgada en los extremos. Los microtúbulos del huso fijan a los cromosomas en sus centrómeros. Cuando está totalmente formado el huso, la envoltura nuclear se desintegra: ésta constituida a su vez por dos membranas, se rompe en vesículas que semejan piezas de retículo endoplásmico. El rompimiento de la envoltura nuclear permite que los microtúbulos del huso invadan la región nuclear. Algunos de éstos microtúbulos se unirán a los cromosomas y algunos interactuarán entre sí. Cada cromátida hermana tiene una estructura llamada cinetocoro, ubicada en el centrómero (véase fig. 29. ). Por lo tanto, cada cromosoma duplicado tiene dos cinetocoros. Cada cinetocoro
se fija en algunos de los microtúbulos cercanos (que ahora reciben
el nombre de microtúbulos cinetocóricos) conforme invaden
la región nuclear. Aunque el mecanismo exacto aun esta bajo investigación,
el cinetocoro de una cromátida hermana de cada cromosoma
En células humanas la profase dura de 30 a 60 minutos.
Durante la metafase (la “etapa intermedia”), los microtúbulos cromosómicos que corren hacia polos opuestos participan en una batalla, en la cual cada uno de ellos jala hacia su propio polo. Parece ser que, los microtúbulos cinetocóricos largos jalan más fuerte que los cortos. Por lo tanto, si un cromosoma está más alejado de un cromosoma que del otro, es arrastrado hacia el polo más distante. También es posible que en ésta etapa, los cromosomas sean alejados de los polos; mientras más cercano se encuentre un cromosoma a un polo, será empujado con mayor fuerza. Al ser empujado hacia el polo más distante o alejado del polo más cercano, o ambas cosas, cada cromosoma termina alineado a lo largo del eje ecuatorial de la célula, el centro entre ambos polos. La metafase termina cuando todos los cromosomas se han alineado en el ecuador, y tiene una duración de 2 a 6 minutos. Durante la anafase, las cromátidas hermanas se separan y son llevadas a polos opuestos de la célula. Al inicio de la anafase, el centrómero de cada cromosoma se divide y las cromátidas hijas se separan en dos cromosomas hijos independientes. Los cinetocoros realizan algunos movimientos moleculares. Los “motores” proteicos dentro de los cinetocoros, parecidos a los que ocasionan la inclinación de los cilios y los flagelos, jalan a los cinetocoros (y a los cromosomas unidos a ellos) hacia el polo a lo largo de los microtúbulos cinetocóricos. Simultáneamente, los microtúbulos se desensamblan dentro de los cinetocoros de tal manera que los microtúbulos se acortan aproximadamente a la misma velocidad que los cinetocoros se mueven hacia los polos. Observe que las cromátidas hermanas de cada cromosoma se mueven hacia cada polo de la célula. Debido a que ellas son copias idénticas de los cromosomas originales, cada uno de los dos juegos de cromosomas son idénticos. La mitosis funciona igual en células que tienen unos cuantos cromosomas, como las de las moscas de la fruta (8 cromosomas), que en las que contienen cientos de ellos, como las células de las “colas de caballo” y de algunos helechos. Esto mismo ocurre en las células haploides, diploides y poliploides. La anafase tiene una duración aproximada de 3 a 15 minutos.
Cuando los cromosomas llegan a los polos, se inicia la telofase (la “etapa final”). El huso se desintegra. Las vesículas que se formaron, cuando se ha roto la antigua membrana nuclear durante la profase tardía, se unen alrededor de cada grupo de cromosomas formando dos nuevas membranas nucleares. Los cromosomas se extienden nuevamente y reaparece el nucleolo. En casi todas las células, la citocinesis ocurre durante la telofase, encerrando cada uno de los núcleos hijos en una célula independiente. Duración aproximada en la telofase de 30 a 60 minutos.
En la mayor parte de las células, durante la telofase se inicia
la división del citoplasma en dos mitades casi iguales. En las
células animales, los microfilamentos compuestos de las proteínas
actina y miosina forman anillos alrededor del plano ecuatorial de la célula,
rodeando los restos del huso mitótico. Los microfilamentos se fijan
a la membrana plasmática. Durante la citocinesis, los anillos se
contraen y jalan el plano ecuatorial de la célula, igual que si
usted jalara la cinta que se encuentra en la cintura en unos pantalones
deportivos. Finalmente, la “cintura” se contrae completamente,
dividiendo el citoplasma en dos células hijas. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
j) Actividad de aprendizaje – Mitosis |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Coloca una cebolla en un vaso de agua de manera que la base
del bulbo quede en contacto con la superficie del líquido. Para sostener
la cebolla puedes clavarle lateralmente cuatro palillos de dientes en forma
de cruz. Al cabo de dos o tres días habrán crecido las raíces
y podrás cortarles la punta o, mejor dicho, el ápice.
Coloca el fragmento cortado en un portaobjetos excavado y, con el propósito de liberar a las células de su cápsula de secreción, agrégale dos o tres gotas de una solución de ácido clorhídrico (5N) durante quince minutos. Si en lugar de esta solución (5N) utilizas una más concentrada, achicharrarás a las células y tendrás que empezar de nuevo. Limpia la preparación con unas gotas de agua destilada. Usándolo a manera de secante, retira el exceso con papel filtro. Agrega dos o tres gotas de acetorceína y espera tres minutos. Si las células resultaron sobreteñidas, ensaya lapsos más breves. Coloca la preparación entre un portaobjetos plano y un cubreobjetos y, presionando sobre éste con la goma de un lápiz, aplasta el fragmento de raíz. Retira el exceso de colorante con papel filtro y aclara con unas gotas de ácido acético al 45%. Calienta una aguja de disección, acércala a un trozo de parafina y, antes de que se enfríe, colócala en la línea de unión entre el portaobjetos y el cubreobjetos. Repite este procedimiento hasta que la preparación quede sellada por los cuatro costados del cubreobjetos y podrás observarla durante un buen rato sin peligro de que se seque. Lo anterior constituye una práctica de la que ningún curso respetable de biología puede prescindir. Pero puede ocurrir que al realizarla resulte que no aparece ninguna célula en división. ¿Cómo es posible que una raíz que te consta que tiene células en división –puesto que la has visto crecer—no muestra más que células en interfase? Conste que la pregunta solamente es válida si has realizado bien la preparación, es decir, si las células pueden distinguirse con claridad. ¿No será entonces que el momento elegido para hacer los cortes no era el adecuado? Si se traduce esta pregunta al estilo ideal para plantear los problemas de acuerdo con el método experimental, se tendrá lo siguiente: ¿cuál es la relación entre la frecuencia de la mitosis y la hora del día? Acaso ya estés harto de que se te diga “consigue, pon, sostén,
corta, coloca, agrega, limpia, calienta, repite y demás”.
Aquí tienes entonces el punto de partida para una pequeña
investigación. Tú decidirás cómo hacerla. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
II. Cuestionario de Autoevaluación |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 1.- Se encarga del estudio de las células a) Fisiología b) Citología c) Anatomía d) Patología 2.- Agregó el tercer postulado a la teoría celular a) Robert Brown b) Matthias Schleiden c) Rudolph Virchow d) Robert Hooke 3.- Según la teoría celular todas las células se derivan de: a) La materia inorgánica b) La materia orgánica c) Cultivos en cajas de petri d) Células preexistentes 4.- Las células fueron denominadas así por: a) Dutrochet b) Hooke c) Schleiden d) Schwann 5.- El limite externo de una célula animal es: a) La membrana plasmática b) El núcleo c) El citoplasma d) Pared celular 6.- ¿Qué organelo contiene el material genético de la célula? a) El aparato de Golgi b) Los Ribosomas c) El núcleo d) Las mitocondrias 7.- La mayor parte del ATP de la célula se sintetiza en: a) El cloroplasto b) Ribosomas c) El núcleo d) Las mitocondrias 8.- ¿Qué organelo clasifica, modifica químicamente y empaca las proteínas recientemente sintetizadas. a) El aparato de Golgi b) Los Ribosomas c) El núcleo d) El cloroplasto 9.- Los organelos rodeados de membrana, que se encargan de la digestión de sustancias y que contienen enzimas reciben el nombre de: a) Lisosomas b) Retículo endoplásmico liso c) Cilios d) Mitocondrias 10.- Un conjunto de canales membranosos, tachonados con ribosomas reciben el nombre de: a) Lisosomas b) Aparato de Golgi c) Retículo endoplásmico rugoso d) Mitocondrias 11.- El transporte activo por la membrana plasmática sucede mediante: a) Difusión b) Proteínas de membrana c) DNA d) Agua 12.- La siguiente es una característica de la membrana
celular |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||