
Paleoecology

* Adaptive functional morphology
CHAPTER 1
Paleoecology is the science which deals with fossils as a tool for detecting ancient environments, in which these fossils lived or supposed to have lived and also the associated sediments were deposited.
This leads us to consider fossils as once living things and not as dry lumps of stones in a museum.
Many sedimentologists try now to interpret depositional environments on the basis of sedimentological features and without the need to fossils, this is not certain in many cases because many sedimentological features can be produced in varying environments and in such cases fossils are the judge!
The greatest precision of environmental interpretation comes from integration of interpretations based on Paleoecology and Sedimentology, as well as on other approaches.
Definition of Paleoecology:
To define Paleoecology we should first define ecology, which is the study of the interactions of organisms with one another and with the physical environment. In such case Paleoecology can be defined as the study of the interactions of organisms with one another and with the physical environment in the geologic past.
This is the simple definition of both ecology and paleoecology. The two definitions seem similar however; application and emphasis of both are distinctly different because:
1. Incomplete preservation of fossil record precludes the examination of many of the standard topics of ecology.
2. Role of time: Ecologists deal with processes taking place during short time span and which are difficult to be observed in the geologic record by paleoecologists who work within a framework of thousands or millions of years. Consequently, both evolutionary processes and long-term environmental change are lacked in ecology.
3. The ecologist is able to examine directly the totality of the ecosystem under study; to determine the life histories and interactions of all the organisms present and to relate these biologic data to the instantaneous characteristics of the physical environment. In contrast, the paleoecologist can't do that because most of the organisms in the fossil record even the most abundant taxa are not preserved.
In general, however, the capabilities of paleoecology, as compared to those of ecology are more encouraging for two reasons:
1. The generality of ecologic models can only be tested in contemporaneous environments but not through time because of the short span of scientific observation. So time factor is most readily available for paleoecologist.
2. Ecologic studies depend on limited components of the biota gathered only periodically and not on comprehensive data as we might think. So, paleoecology and ecology are based on similar information either in quality and quantity.
In summary, paleoecology is not a mere tool to understand the physical aspects of environments but it has many other applications such as:
1. Understanding fossils as once-living organisms.
2. Understanding fossils as once-integrated communities and ecosystems.
3. Understanding range charts, which represent biogeographic changes through geologic time.
DATA BASE IN PALEOECOLOGY:
Paleoenvironmental reconstruction depends on three ingredients:
1. A well-established stratigraphic framework.
The stratigraphic setting provides the spatial and temporal relationships for the comparisons of fossils within geologic history.
2. Good taxonomy.
Good taxonomy of fossils, which are the basic data of paleoecology means they should be adequately identified and correctly positioned within the stratigraphic framework.
3. A comprehensive ecologic background.
Ecologic background consists of an understanding the ways in which living organisms function within their ecosystem: how their morphology and physiology is adaptive to their conditions of life, the ways in which they may interact with one another, and the ways in which they may modify their life history to fit the environment.
PALEOECOLOGY AND BIOLOGY:
Biology is the science of studying living animals (Zoology) and plants (botany). The relationship between the organisms and their environment today is the science of Ecology. So, ecology tries to answer how and where organisms live today. Paleoecology on the other hand tries to answer the same question but on fossil organisms.


The modern ecologist studies a living population like this year's automobile - as a working machine. The paleoecologist studies a whole parking lot full of abandoned earlier models extending back to the horsless carriage. All stationary, but containing within them the evidence of how this year's model came to be what it is. The ecologist's automobile is dynamic in space but static in time. The paleoecologist's automobiles are static in space but dynamic in time.
PALEOECOLOGY AND PALEONTOLOGY:
As we said before, biology is the science of studying living organisms, paleontology is on the other hand, the science of studying ancient organisms (fossils).
There is a great difficulty in studying the ecology of fossils because the physiological activities of these fossils have long since ceased, and no one has yet found a fossilized chromosome.
What should we do then? There is no way other than careful examination and description of fossils from their morphology. It is advised that a paleoecologist should have good paleontologic bases and should revise his fossils carefully to get accurate paleoecologic results. He shouldn't think that he can just sample the cream and leave the milk to someone else. The relation of paleoecology to paleontology lies in the matter of evolution. The doctrine of the survival of the fittest is an ecological matter, since those fittest to survive are those best adapted in terms of habit and habitat.
PALEOECOLOGY AND GEOLOGY:
Paleoecology is an important science in serving geology especially in determining the environment of deposition represented in the rock.
There are applied paleoecology which is concerned with what fossils can tell about the conditions under which the sediments were deposited, and pure paleoecology which is concerned with what sediments can tell about the conditions under which fossils lived.
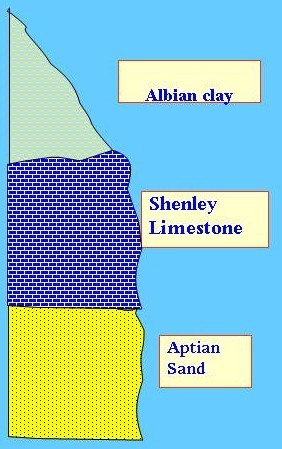 So,
consideration of environment and its effect on fauna is very important. An
example for this is the Shenley Limestone in the Cretaceous of England, which
consists of highly fossiliferous limestone lenticles above Aptian sand and
Albian clay. Because the fossils found in this limestone are apparently similar
to Cenomanian fossils, the Shenley L.S. was dated as Cenomanian and its presence
below Albian was interpreted as due to overturning during Pleistocene glaciation.
Later studies showed that the fossils are in fact Early Albian and its
similarity to Cenomanian is just environmental. In this case the section is
normal and no overturning took place!
So,
consideration of environment and its effect on fauna is very important. An
example for this is the Shenley Limestone in the Cretaceous of England, which
consists of highly fossiliferous limestone lenticles above Aptian sand and
Albian clay. Because the fossils found in this limestone are apparently similar
to Cenomanian fossils, the Shenley L.S. was dated as Cenomanian and its presence
below Albian was interpreted as due to overturning during Pleistocene glaciation.
Later studies showed that the fossils are in fact Early Albian and its
similarity to Cenomanian is just environmental. In this case the section is
normal and no overturning took place!
ADAPTATION TO ENVIRONMENT:
v Every organism adapts itself to the environment in which he lives and which is either wide or narrow.
v Examples of organisms which have limited habitat are the deers of the national forest south of the Grand Canyon. The killing of predatory animals led to great increase in the numbers of the deers. This led to great depredation on the local plants and this caused mass winter starvation for the deers until they were reduced to a tenth of their peak population.
v On the other hand, there are other forms which tolerate a wide range of environment. The most tolerant of all species at the present day is MAN. He can live in the hottest regions as well as the coldest regions, he can live in villages and also in modern cities, he depends utterly for his food on people he never sees and who may live on the other side of the world.
ADAPTATION TO MODE OF LIFE:
Every organism is adapted to a certain mode of life. Thus many different animals resemble each other superficially because of adaptation to the same habit (swimming for example, Fig. 1, 2). On the contrary; closely related forms may differ from each other because of adaptation to different habit. For example sponges tend to develop into irregular encrusting forms in strong currents, but into tall symmetrical shapes in quieter waters.
APPLICATION OF PALEOECOLOGY
Paleoecology has certain applications in oil prospection especially when oil is found associated with reefs or in interdigitating marine and non-marine deposits.
* In case when oil is associated with reefs we should do the following:
1. Delineation of the shoals which acted as foundations for the reefs by finding the highest calcarenite ratio.
2. Delineation of the reefs themselves by finding the highest ratio of potential frame-building organisms (grainstone or boundstone).
3. Delineation of reef fronts by study of the distribution patterns of the different organisms and their comparison with known reefs.
* In case when oil is accumulated in interdigitating marine and non-marine deposits, quantitative studies of the ecological distribution of foraminifera are particularly useful in predicting the proximity of facies margins.
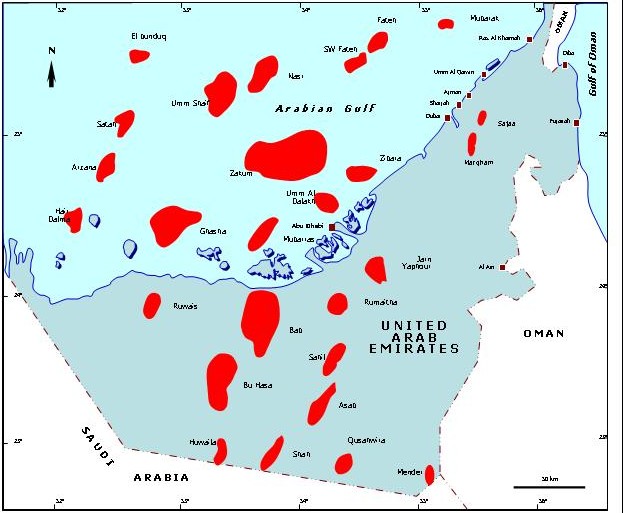
An example of applying paleoecology in oil prospecting
Abu Dhabi. It is situated 85 Km W-NW of Asab Field (Fig. 1) and produces oil from the Shuaiba Formation (Aptian).
Sediments:
The sedimentary composition of Shuaiba Formation is represented by reefal limestone, which consists of skeletons of frame-building organisms, mainly rudists with some corals, algae, large forams (Orbitolina), ammonites and planktonic forams.
The microfacies study allowed the recognition of nine lithofacies types:
1. Whole rudist and large rudist fragments in a lime mud matrix, sometimes pellety and intraclastic. Hardground present.
2. Grainstone and packstone predominantly composed of rudist debris.
3. Lithocodium Boundstone: heterogeneous sediments bound by Lithocodium colonies.
4. Bacinella Boundstone: Miliolid Wackstones and packstones and pellet grainstone, bound by Bacinella.
5. Wackstones and occasional packstone with common Orbitolina.
6. Chalky lime mudstone with occasional planktonic foraminifera and clacispheres.
7. Dense micrograined, cemented lime mudstones with globigerinids and ammonites.
8. Micrograined cemented lime mudstones and wackstones.
9. Heterogeneous wackstones and packstones with lime mud matrix: skeletal elements include miliolids, orbitolina, small rudists, algal debris and pellets. Hardgrounds present.
Paleoenvironmental History:
From the lithofacies and biofacies aspects, it seems that the Shuaiba Formation underwent three depositional phases:
Phase1: dominated by algal growth (Bacinella) reflecting shallow open marine environment of subdued topography. Slight thickening is observed at the south. The succeeding limemuds with variable fauna of globigerinids, Orbitolina, echinoid and molluscan debris reflect a southward transgression of deeper marine conditions.
Phase2: the conditions became shallow again over southern Bu Hasa, allowing the accumulation of more than 200 ft of algal boundstones with some corals. In the northern Bu Hasa, slightly deeper conditions retarded algal growth and allowed the deposition of argillaceous limemud with globigerinids. The thickness of algal sediments decreases to 40 ft. and then to 15 ft. further north.
Phase3: Rudistids colonized the previously formed algal platform in the most elevated parts, it was colonized by caprotinids with fewer caprinids. The depressions and channels on the platform remained too deep for rudist colonization and instead argillaceous limemud rich in Orbitolina were deposited.
To the north bioclastic sediments accumulated and provided a protecting shoal system behind which the rudist colonies were able to thrive in shallow quiet water conditions.
Late Shuaiba regressive sedimentation caused the rudist reef to migrate northward over the deeper sediments.
Summary:
Core samples and well log data obtained form the wells drilled in Bu Hasa field allowed the recognition of a well developed rudist reef in the area. This reef is divided into three zones:
- A zone of maximum building which has the maximum thickness and from which most oil is produced (Fig.2 & 3).
- An outer buildup zone lies to the northeast of the maximum (fore reef) and
- An inner buildup zone lies to the southwest of the maximum (back reef).
Clearly the reef was affected by diagenetic process which controlled its quality as a reservoir rock. Some of these processes had a positive effect in improving reservoir permeability such as solution and compaction fracturing. Others had a negative effect such as pressure solution and stylolithification associated with depth of burial, this reduced pore volume and so hindering cross flow within and between reservoir units.
COAL:
Paleoecology has also direct applications in coal exploration especially in determining coal-Bearing strata and their lateral thickness variation.
An example for that is the paleontologic and paleoecologic study to correlate the cyclothemes of Europe. There we find marine bands of Upper Carboniferous coal cycles rich in faunas of Goniatites, crinoids, brachiopods, pelecypods and the rest are often represented laterally by bands rich in Lingula. Although the same horizon but in a less marine environment (brackish).
Uniformitarianism means present is the key to the past, so we use what is going on today to interpret past geologic events.
Uniformitarianism is divided into two types:
1. Substantive uniformitarianism:
This implies that: materials, conditions and rates of processes during earth history have remained constant.
2- Methodological uniformitarianism:
This implies that the laws of nature (such as gravity, properties of fluid flow, and thermodynamics have been constant in their operation through geologic time.
Of course earth materials have not remained constant in composition and proportions during geologic time, and the rates of some processes fluctuated widely beyond the ranges presently observed. Thus the present may not be a good key to the past and substantive uniformitarianism should be abandoned.
On the other hand, laws of nature haven't changed and are applied now in many fields, for example, thermodynamic principles can be applied in experimental petrology to generate phase diagrams that specify the physical and chemical conditions of igneous and metamorphic phenomena. Also hydrodynamic laws can be applied to understand the fluid characteristics resulting in the deposition of sediment with specific textural features and sedimentary structures.
Paleoecology and Uniformitarianism:
In paleoecology, the laws of nature can't be easily used to detect ancient environments but we depend on the habitat characteristics of the most closely related living taxon. In this case we use substantive rather than methodological uniformitarianism. And since it depends on taxonomic comparisons, it is therefor called taxonomic uniformitarianism. Functional morphology and oxygen isotopic paleo-temperature determination methods are of the methodological uniformitarianism.
Examples of applying taxonomic uniformitarianism:
The taxonomic uniformitarian approach can be
applied at many different taxonomic levels. Ex. A Pleistocene limestone may
contain specimens of the coral Acropora palmata. This species is living
today in the Caribbean area at very shallow (<5m), turbulent water in the open
reef environment. It is most abundant on the shallowest, most turbulent portion
of the reef. Thus the depositional environment of the limestone containing this
species (assuming that the fossil is in place) can be determined by
uniformitarian approach.

Another limestone of Miocene age may contain a
now extinct species of Acropora. The entire genus Acropora of
course has a much broader range of environmental requirements than any one of
its species. The genus is widespread in reef and associated environments
throughout the world. Two common species of Acropora occur in the
Caribbean area, A. palmata and A. cervicornis. A.
cervicornis is most abundant in moderately turbulent water in the back reef
area and on patch reefs some distance from the shelf edge in less turbulent
environments than A. palmata. The total environmental range would be
further broadened if the many Indo-pacific species are included. Thus: the
taxonomic uniformitarian approach used at the generic level would give less
precise information than at the species level, but would still indicate a
probable reef or reef-associated environment. x
At the family level, we will get less precise information. Ex. An early Cenozoic limestone may contain Dendracis, an extinct genus of the family Acroporidae. This family consists largely of species of the genus Acropora but includes a few other less common genera. All live in tropical, shallow, full salinity environments, but their total environmental range is greater than that for the genus Acropora and certainly for any species.
A Cretaceous limestone may contain a species of an extinct Scleractinian family such as Stylinidae. The order Scleractinia includes all of the modern hexacorals from the reef building forms of the tropics to the relatively deep water, ahermatypic forms of the high latitudes. The total environmental range of the order is much broader than lower taxonomic levels. Most are tropical. Most live in shallow water. All live in normal or near normal salinity. All require well to moderately well oxygenated water that is relatively sediment free.
So the presence of a fossil Scleractinia gives much less specific information although they generally suggest shallow tropical seas. A Paleozoic limestone may contain an extinct order, Rugosa (class Anthozoa). Modern Anthozoans include the orders Octacorallia and Scleractinia and several other orders without skeletons. Modern anthozoans are most abundant in the shallow tropics but are common down to considerable depths. They occur at temperatures from o to 40ºC or above. They are most common at normal marine salinity but some forms extend into brackish or hypersaline waters. Consequently a very limited amount of information concerning environment can be obtained by taxonomic uniformitarianism at the class level. Even less could be said at the phylum level. Coelenterates occur in particularly all aquatic environments, both marine and nonmarine. So, Substantive taxonomic uniformitarianism increases in precision with decreasing geologic age and the best results are obtained in rocks of Late Cenozoic age which contain many extant species or at least genera. On the other hand, methodological uniformitarian approach (functional morphology) can be used for older as well as younger species.
Role of differential preservation in applying taxonomic uniformitarianism
Differential preservation may cause the final assemblage investigated by the paleoecologist to be quite different from the original living assemblage, in this case the assemblage will give another different environment.
Ex. In the Upper Cretaceous rocks of New Jersey, the foraminiferal assemblages consist entirely of planktonic species (deep environment). Modern assemblages dominated by planktonic species are almost exclusively found in deep sediments, although planktonic species also occur in shallow water sediments along with more abundant benthonic species.
Applying the uniformitarian approach, these Upper Cretaceous rocks were deposited in deep water. On the contrary, these rocks contain sedimentary features suggesting deposition in shallow water.
More important from the paleoecologic point of view, the total fossil assemblage also contains oysters, which by the uniformitarian approach suggest shallow water.
What is the reason?
The reason is that the assemblage contains only fossils that were originally composed of low Mg-calcite. Aragonitic bivalves and gastropods which were originally in the assemblage were gone. Benthonic foraminifera composed of high Mg-calcite also dissolved.
How can we use taxonomic uniformitarianism in detecting or studying ancient environmental parameters?
Environmental parameters include currents and water turbulence, temperature, salinity, nutrient elements (productivity), dissolved oxygen levels, depth and substrate type.
Currents
There are two types of currents:
1- Oceanic currents (global circulation).
2- Local currents (generated by waves and tides).
Global circulation (horizontal and vertical):
Global currents are very important in Paleoecology, their effect on organisms is indirect as they control other environmental parameters such as temperature, salinity and concentration of nutrients. They may also directly affect distribution of biota by transporting pelagic species, transporting the pelagic larval stage of many benthonic forms and in some cases even transporting shells after death of the organism (e.g. Nautilus)
Wind is the driving force behind surface (horizontal) currents. The oceanic circulation pattern is directly related to the pattern of global winds and pressure system.
On the other hand, vertical circulation is more important. It is controlled by density differences in the water. Factors controlling the density of seawater are temperature, salinity and suspended sediment content. Temperature is the most important of these. At low latitudes where the temperature difference between the surface and bottom are greatest, the oceans are density stratified with the warmer, less dense water on top and cooler, more dense water on the bottom. Vertical circulation is thus weak. At high latitudes, cooling of seawater produce dense water, there is no density stratification as in low Latitudes and hence vertical circulation is strong. This dense water sinks to the bottom and slowly flows along the bottom of all the oceans (Fig. 2.6). This deep water slowly warms due to heat flow through the sea floor and gradual mixing with the less dense water above. It will eventually return to the surface.
1- Recycling nutrient elements released by decay of organic matter, which sinks to the bottom.
2- Brings dissolved oxygen from the surface to the depths where it can be utilized by organisms living there.
3- Occurrence of divergent and convergent currents. When surface currents diverge from one another or move away from land areas, water must rise from below to replace the water that has moved away laterally. This process is called upwelling. The water that “wells up” from below is richer in nutrient elements than the water it replaces.
Local currents are generated by winds, tides and local density differences (e.g. turbidity currents), or they may be local expressions of global currents.
Winds:
Wind-generated currents are usually associated with waves. The motion of waves causes water below to move in closed orbits (Fig. 2.8a), the diameter of the orbit decreases with depth until motion ceases. Thus wave-related currents are effective only in shallow water. When waves break, water is piled up along the shore. If the waves do not break perfectly perpendicular to shore, they will move parallel to the shoreline causing the formation of long shore currents. These long-shore currents must return offshore causing the formation of another kind of currents known as Rip currents (Fig. 2.9).
Tides:
Tides also generate currents but their effects are limited to shallow water.
Density-difference:
Density-generated currents, especially turbidity currents, are very important in their effects on organisms. They cause burial of fossils and change substrate nature. They are effective in shallow as well as deep water.
Strong currents have a number of advantages to organisms such as:
1- Bring food to the organism.
2- Remove waste products.
3- Bring dissolved oxygen.
4- Bring nutrient elements for plants.
5- May eliminate predators that are not able to cope with life in strong currents.
Organisms can’t hold themselves or maintain position, so they adopt a number of strategies to cope with this problem as follow:
Sessile organisms: develop strong attachment (Table 2.2) by:
1) Cementation 2) byssus 3) flexible or articulated skeletons
4) Strong muscle attachment 5) suction or hold fasts of various types.
· Branching organisms such as some corals orient their branches in the direction of strong currents (Fig. 2.10). Such orientation can be used to determine paleocurrent strength and direction.
Vagile organisms: develop strong locomotion (certain bivalves and gastropods). Or:
· Live in burrows to escape the strong currents, but extend siphons or other food-collecting devices into the current (burrowing bivalves).
· Develop streamlining or a low profile to minimize the drag in currents (limpets and chiton).
On the other hand, life in weak currents has a number of advantages such as:
1- No energy loss in adaptation with strong currents.
2- Energy is devoted to food gathering and other life processes.
3- More variability in morphology.
Disadvantages of weak currents:
The organisms must produce their own currents to bring in food and dissolved oxygen and to eliminate wastes.
Ex. Sponge produces currents by flagellated cells to get the suspended organic matters. Many other organisms are deposit- feeders.
Other Effects of currents:
Currents also determine the substrate kind, Hard or coarse substrates are more common in high currents, soft substrates in low currents.
High currents are generally confined to shallow environments whereas quiet water may be either shallow or deep.
Temperature
The temperature range in the ocean is approximately –2 - 40ºC. Coldest waters are found in the Polar Regions, where seawater is freezing. Highest surface temperatures occur in lagoons or bay settings marginal to tropical oceans, where the temperature may sometimes exceed 40ºC. Much higher temperatures recently have been recorded in thermal springs associated with submarine rift zones. In non-marine environments, the temperature range is much greater, ranging from about –50ºC to nearly 100ºC in some continental hot springs.
Vertical Variation:
In Polar Regions, bottom water has the coldest surface water temperature, this is due to cooling and sinking of surface water.
Geographic Variation:
Surface temperature is controlled by solar energy. It varies inversely with latitudes however, in areas with upwelling currents, temperature is low irrespective of where they are. For example the extensive band of slightly lower temperature near the equator in the Pacific Ocean. In nonmarine environments, temperature is more variable and is controlled by altitude and nearness to large water bodies.
Biological Effect:
Temperature has a great effect on metabolic rate, this rate increases with increasing temp. At higher temperatures, metabolic rate decreases and death ensue. However in some cases, organisms can change their metabolic rate with changing temperature, i.e. Those in warm environment have slower metabolic rate than those in cold environment.
In fossil specimens it is difficult to measure metabolic rate, but this is expressed by growth rate. Growth rate can be recorded in fossils, for example the growth rings in the shells. Such rings allow us to determine the average annual growth rate which maybe a function of average annual temperature.
Temperature also controls the time of reproduction. It also determines the geographic range of a species. Temperature may also influence the size of the organism, for example species living in warm water (supersaturated with CaCO3) grow large skeletons.
Salinity
Salinity is a measure of the concentration of dissolved salts in seawater. It is defined as the weight in grams of dissolved solids in 1K of seawater. Salinity is expressed in terms of per mille %o which is equivalent to part per thousand or ppt.
The most common cations in seawater are Na+, Mg2+, Ca2+, k+ and Sr2+. The most common anions are Cl-, SO42-, Br- and F-. The relative proportions of these elements in seawater are constant regardless of absolute concentration. For this reason, they are called conservative elements.
Other elements such as P, N and Si are low and highly variable in concentration due to involvement in photosynthesis and biological processes. These elements are called nutrient elements.
Normal seawater has a salinity of approximately 35 %o. Salinity may be much higher in evaporitic settings such as restricted lagoons and Salinas. Salinity decreases to near zero in areas of mixing with fresh water. It is lowest in high latitudes where evaporation is minimal and some runoff of fresh water from the continents occurs.
Biological Effect:
The concentrations of dissolved solids in seawater (salinity) are very important to organisms. Some organisms are very tolerant of Salinity variation (euryhaline), others have narrow tolerances (stenohaline).
Salinity affects the osmotic pressure across cell membranes within organism. Some organisms can regulate ionic concentration within their body (osmoregulators), but others do not (osmoconformers). In addition to that, some organisms can migrate when local salinity becomes unfavorable, others can close themselves in their shells until favorable salinity returns. Burrowing forms may withdraw deep within their burrows during times of unfavorable salinity. The greatest diversity of marine invertebrates is in area of normal salinity. Diversity declines at both lower and higher salinity but increase somewhat in fresh water (Fig. 2. 19), probably due to stability of salinity in fresh water (essentially 0). Saturation of water with CaCO3 also affects the thickness of the shell, the thickness increases with the increase in CaCO3 concentration.
Nutrient Elements
Nutrient elements together with light are very important in determining the quantity of organisms living in a given environment (marine). In nonmarine (terrestrial) environment, temperature and availability of water are additional factors.
Productivity:
Productivity is the rate of production of organic material by biological processes. It is divided into primary productivity (the production of organic material by photosynthesis) and secondary productivity (The production of organic material by animals utilizing preexisting plant and animal organic matter).
Biomass (Standing Crop):
This is the amount of living organic material present in a given area at any one time. It is related to but not equivalent to productivity. Biomass is a function of both longevity as well as productivity. For example: an area with large long-lived plants will have a larger Biomass than an equally productive area with small, short-lived plants.
How can we measure productivity or Biomass in the geologic record?
In fact this is difficult and uncertain. But knowing something about productivity is so important to full understanding of Paleoecology.
Fossil Biomass is difficult to measure for two reasons:
1. Very little of the original Biomass is preserved in the geologic record.
2. The concentration of fossils is controlled not only by the rate of their production but also by sedimentation rate.
Since Biomass refers to the organic portion of organisms which is almost never preserved, The concentration of skeletal materials may be crudely correlated with Biomass under certain condition (non-preservation of soft-bodied organisms). Even the concentration of fossilized skeletal materials is subject to diagenetic factors and may not accurately reflect the original concentration.
Productivity is also difficult to measure because longevity of the organisms involved is not known.
Vertical Variation:
Light is the energy source for the formation of organic material by photosynthesis. It is always available in shallow water, but its distribution may be seasonal. On the other hand, concentration of the nutrient element, N, P, is the limiting factor.
With increasing depth, the availability of light as an energy source becomes more important than nutrient element concentration.
At shallow depth plants are photosynthesizing faster than they are respiring, thus more organic material is being produced than is being consumed. At the compensation depth photosynthesis and respiration are equal but below it respiration exceeds photosynthesis and thus plants can't survive.
Destruction of organic matter frees nutrient elements to go back into solution in the seawater. Also some organisms produce organic material using chemical energy rather than light energy, for example bacteria which obtain their energy by oxidizing sulphides in the thermal spring associated with spreading centers.
Geographic variation:
Geographic variation in nutrient element concentration and productivity is strongly influenced by global circulation patterns. Areas of divergent current are usually areas of high nutrient element concentration and of high productivity. This is because water enriched in nutrient element from below the compensation depth upwells in these areas.
Productivity is on average higher near continent than in the centers of oceans due to three factors:
1- Upwelling often occurs near coasts where surface water move away from the continent.
2- Increase of nutrient element concentration brought by rivers.
3- Organic material produced by photosynthesis accumulates on the shallow continental shelf.
Q. Which areas are more productive, surface water in high latitudes (Polar regions) or that in low latitudes Tropical regions)?
Biological effects:
Surface water productivity which depends on nutrient elements concentration affects the nature of organisms found in a given area.
Hermatypic corals are adapted for life in areas of low productivity, this is because when productivity increases, other organisms tend to replace corals.
Productivity also influence depth zonation of light dependent organisms such as hermatypic corals and algae. These organisms will vary in their depth distributions as productivity levels vary (Fig. 2.22).
Depth
Depth at which sediments were deposited is always the question of geologists. Water depth is important in interpreting paleogeography and geologic history. It is also important in studying sedimentary basins and the occurrence of hydrocarbons or other economic resources within these basins.
Water depth also influences sedimentary process and properties. For this reason Sedimentologists determine water depth by sedimentologic methods without the need to paleoecology. This is not always definitive. For example very fine grained sediments (shale) may be especially difficult to interpret, they were clearly deposited under quiet-water conditions, but was the water quiet, because it was deep or because it was protected by some barriers from wave action?
Water depth affects many other factors such as hydrostatic pressure, light, temperature, salinity, and concentration of nutrient elements and quantity of dissolved gases.
Variation in distribution of organisms such as foraminifera is shown in fig (2.23).
Light factor can be used in determining paleodepths. It is necessary for photosynthesis, so the abundance and characteristics of in-situ benthonic plants correlates strongly with depth. Plant abundance decreases rapidly in the lower part of the photic zone and plants are absent below. Some plants such as the red algae are able to photosynthesis at depth 250 m, others such as green algae have a much shallower limit.
Many animal groups have genera that contain symbiotic algae within their tissues (corals, foraminifera, bivalves) these are strongly controlled by light and depth.
Therefore, we can conclude that depth affects distribution and morphology of plants as well as invertebrate animals (table 2.5).
Dissolved oxygen
Dissolved oxygen is very important in the distribution of organisms, it also determines early diagenetic processes at the sediment-water interface or at shallow depths within the sediment (oxic or anoxic).
Oxic (oxidizing) Hematite, Carbonates
Anoxic (reducing) Pyrite , Sulphides
It also affects preservation of organic compounds
Plants No O2 Coal
Animals No O2 Kerogen or oil
Organic content of sediments has major economic
implications in terms of hydrocarbon source beds. Surface water is rich in
oxygen dissolved from the atmosphere or from photosynthesis. Oxygen levels
decline with depth especially if there is no vertical circulation.
Oxygen levels are low at very shallow depths in restricted shallow seas, lakes, swamps and marshes, where oxidation of very abundant organic matter removes oxygen.
Oxygen levels are balanced by the interplay of rate of production of oxygen by photosynthesis, depletion by respiration and oxidation and vertical circulation.
During Middle Cretaceous, anaerobic conditions prevailed. This may have been due to:
1- Sluggish circulation due to a low temperature gradient between poles and equator or salinity stratification due to higher salinity water at depth than at the surface.
2- High productivity, although sluggish vertical circulation leads to low productivity due to slow recycling of nutrient elements.
Biological Effect:
1- Fewer organisms are able to survive as oxygen decreases.
2- Some organisms such as hermatypic corals and cephalopod require relativity high oxygen levels. Other such as some annelids (worms) do well in low oxygen environment.
Substrate
There are two types of substrate, soft substrate and hard substrate, the nature of the substrate is determined by many factors such as current strength, sediment source, sedimentation processes and rates, diagenetic processes and the organisms themselves.
Hard ground is a kind of hard substrate consisting of lithified sediments. Lithification may occur after burial, in this case erosion (unconformity formation) is necessary to expose the hard ground later at the surface, hard grounds can also be produced by submarine cementation at the sediment water interface, especially in carbonates. Exoskeleton of organisms themselves may serve directly as hard substrates.
Filamentous algae may add firmness to the substrate by forming mat-like surface.
Organisms adapt themselves to the kind of substrate. In case of soft substrates with rapid sedimentation rate, organisms adapt themselves to keep from becoming buried or to keep the food gathering structures from becoming clogged with sediment. Rapid vertical growth and flexible articulated skeletons are adaptations for rapid sedimentation rate. Organisms such as brachiopods develop large and flat area of shell next to substrate (snowshoe) and may have large portion of shell buried with commissure floating above the sediment (iceberg). In case of hard substrates, organisms have another way of adaptation. Burrowing forms must develop strong muscles, streamlined shells and surface sculptures in order to penetrate easily. Many organisms attach firmly to the substrate by cementation, byssus, muscle attachment or suction. Some organisms nestle in crevices, or bore into the substrate for attachment and stability. Vagile organisms develop strong locomotion and ability to cling to the substrate in the turbulent water that is often associated with these substrates (Table 2. 6).
=============================================================
CHAPTER 2
Biogeochemistry
Biogeochemistry is the use of chemical properties of fossils to determine paleoenvironmental conditions. These chemical properties include mineral composition of fossil skeletons, their trace and minor element chemistry, their oxygen and carbon isotopic composition as well as geochemistry of preserved organic compounds in the rock record. Such use of chemical properties of fossils depends on the fact that the chemistry of the environment in which skeletons form has a strong influence on the chemistry of the skeleton itself. We have to take in consideration the negative effect of diagenesis on the chemical properties of the fossils, which limits the usefulness of the technique particularly in older fossils.
Paleoecologists try to interpret the environment on the basis of the observed morphologic features only. The unseen chemical attributes of the fossil are in fact an important aspect of that fossil and can tell the paleontologist as much about certain features of the environment in which the fossil lived as can the fossils morphologic features. The most complete possible description of the fossil and its environment would include chemical as well as morphological data.
Factors controlling the chemical properties of fossils:
1- Physical chemical factors
2- Environmental factors
3- Physiologic (genetic) factors
4- Diagenetic factors
The mineral composition of skeletal material is of considerable interest to Paleoecologists both as tool for paleoenvironmental analysis and because of its effect on the preservation of fossils.
Mineral Component:
Many different minerals are found in invertebrate skeletons (Table ). The most important minerals among the invertebrates are the two calcium carbonate polymorphs, calcite and aragonite. Opaline silicates and phosphates also make major contributions. In addition, several less common minerals have also been described.


There are also minor minerals, which serve a particular function. Ex. The chitons have magnetite capping for their radular teeth. This mineral is much harder than the aragonite that makes up the basic skeleton of the chitons. Some echinoids have dolomite in their teeth. Dolomite is also slightly harder than the calcite that makes up the majority of the echinoid test.
The mineralogy of now-extinct groups cannot always be determined with certainty. Some fossils that were originally aragonite may have been converted to calcite. Ex. Rugose corals, tabulates and stromatoporoids, some workers suggested that they were originally aragonite as are the scleractinia. However, details of their skeletal structure suggest that no major diagenetic alteration has occurred.
On the other hand, few Paleozoic organisms (nautiloids, bivalves and gastropods) are still having the aragonite skeleton although the associated tabulate and rugose corals are calcite.
Q. Why is CaCO3 the most common skeletal material?
Factors controlling skeletal mineralogy:
1- Shallow seawater is saturated or supersaturated with CaCO3.
2- Precipitation of CaCO3 is more readily controlled biologically than most minerals.
3- Concentration of CO32- depends on pH & CO2 concentration. Metabolic processes readily affect these parameters.
4- Concentration of dissolved Si²+ and PO4³‾, which are used by some organisms, is much lower than Ca²+.
5- Organisms require strong biological intervention to precipitate opaline silica or phosphates.
Calcium carbonate polymorphs:
Calcium carbonate occurs in skeletons in one of the three polymorphous forms: Calcite, aragonite or vaterite, the latter is rare. Calcite is more stable than aragonite under surface temperature-pressure condition. This instability of aragonite is due to its slightly larger heat of formation than calcite.
On the other hand, aragonite is extremely common in modern marine sediments! The reason for that is the high concentration of Mg in seawater. Aragonite is not unstable in the marine environment and Precipitation of calcite is in the form of high Mg-calcite (>8.5 % MgCO3). This high Mg-calcite is more soluble (less stable) than aragonite. Low Mg-calcite (<8.5 % MgCO3) can't precipitate in full marine seawater under tropical or subtropical temperatures due to the abundance of Mg in seawater. Consequently aragonite and high Mg-calcite form instead. Another reason for the abundance of aragonite is that Mg²+ acts as a crystal poison, inhibiting growth of calcite relative to aragonite.
Genetic control of skeletal mineralogy:
The mineral composition of skeletal material is also partly genetically controlled. This is indicated by the presence of different minerals in different parts of a single organism.
Mineral composition can change during the life of the individual. Ex. Crassostrea (bivalve) is entirely composed of aragonite in the larval shell but the adult shell is almost entirely calcite.
Long-term changes in mineralogy are also found, such as coral, which changed from calcite in the Paleozoic to aragonite today. Primitive gastropods were calcite whereas the more advanced forms are mainly aragonite. Generally, aragonitic groups increase in abundance after the Paleozoic.
Diagenetic Effect on skeletal mineralogy:
Groundwater commonly dissolves mineral constituents of fossils. This led to preservation of a small portion of the skeletal material originally buried in sediments. The evidence for that is the great volume of marine sedimentary rocks that don’t contain fossils.
Why does groundwater dissolve skeleton of fossils?
Because it is undersaturated with respect to the mineral of which the fossil is composed. Also permeable rocks and presence of fossil in a matrix that differ in mineralogy from the fossil facilitates dissolution.
As discussed before, aragonite fossils are rare in ancient rocks due to its high solubility. However, some examples are found where fossils were buried in fine shale or in asphalt (far from groundwater contact).
If the dissolution of aragonite fossil occurs before lithification, the resulting void will collapse and all traces of the fossil may be lost, with the possible exception of an imprint.
If lithification has already occurred before the fossil dissolves, a mold will be preserved, and if the mold is later filled with another mineral, a cast of the original fossil will be preserved.
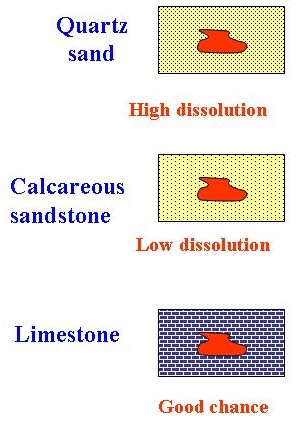
At shallow depths, the seawater is saturated or supersaturated with CaCO3 (except in Polar Regions), so carbonate skeletons are not dissolved. With increasing depth, the oceans become undersaturated with CaCO3 and so dissolution occurs. The depth at which aragonite dissolves is less (500- 2500 m) than that of calcite (3500- 5000 m).
Dissolution is also affected by shell thickness where robust shells suffer less dissolution than delicate ones.
The oceans are everywhere undersaturated with respect to opaline silica, so siliceous skeletons dissolve as soon as they are exposed to seawater. As in the case of carbonate skeletons, the heaviest, most robust forms are most likely to be preserved. Solution of opal continues within the sediment, so that unless sedimentation is rapid, complete loss of opaline fossils is likely.
Geologic application of skeletal mineralogy:
1. Evaluation the degree of preservation of fossil assemblage. Ex. New Jersey Cretaceous fossils, which give different environment due to differential preservation (removal of aragonite shells (.
2. Studying origin and diagenesis of carbonate sediments and rocks, which are composed mainly of fossil skeletons. The relative proportion of calcite and aragonite as well as the amount of Mg in calcite has a large bearing on the way the sediment behaves during lithification. The solution of aragonitic skeletal grains can result in the development of secondary porosity and hence development of oil reservoirs (see Bu Hasa Oil Field).
3. Paleotemperature can’t be successfully determined because:
- The mineralogy-temperature effect differs from species to species and perhaps within genetic variants of a single species. i.e. Aragonite proportion may correspond to a given temperature in one species and to another temperature in another.
- Several other factors affect skeletal mineralogy.
- Finding well-preserved aragonitic specimens is difficult.
---------------------------------------------------------------------------------------------------------------
Trace chemistry deals with studying trace (minor) elements associated with the major elements forming fossil skeletons. The ions of such elements may be present in solid solution in the crystal lattice, absorbed to the crystal surface, incorporated in the organic matrix or incorporated as separate mineral phases.
The concentration of trace elements in a fossil is a function of the physical chemistry of the skeletal formation process, of environmental variables, of the physiology of the organism and of the diagenetic processes.
The most abundant trace elements are Mg and Sr in addition to other elements such as Cd and Mn.
Why are Mg and Sr abundant?
1- The ionic radii of Mg2+ and Sr2+ are close enough to that of Ca2+ to allow ready substitution in calcite or aragonite crystal lattice.
2- The ionic charge in the same as that of Ca2+, facilitating substitution.
3- Mg2+ and to a somewhat lesser extent Sr2+ are abundant in natural water, especially seawater.
Factors controlling trace chemistry:
Molar concentration Molar concentration
\[Me/ Ca] skeleton = K[Me/Ca] water
Where K is the distribution coefficient and depends on temperature (Fig. 3-7).
So: 1- Knowing Me/Ca in a fossil and K for that ratio, the [Me/Ca] water can be determined and so we can get information about environmental conditions and depositional environment.
2- K is not constant but is temperature dependent for some ions.
Thus: Knowing [Me/Ca] for fossil and water, we can determine K. And from the relation between K and T, the temperature of the formation can be determined.
This approach has been used extensively in igneous and metamorphic petrology to determine temperatures of mineral formation.
Limititions:
1- K depends on the mineral form of CaCO3 (calcite or aragonite).
e. g. Mg/ Ca ratio is higher in calcite than in aragonite skeletons.
Conversely: Sr/ Ca ratio is higher in aragonite than in calcite skeletons. (this depends on the ionic radii of Mg and Sr for substitution in the crystal lattice of both minerals, Mg2+ < Sr2+.
2- K is controlled biologically (depends on the physiology of skeletal formation process. This is indicated by:
- Differences between different groups of organisms growing under the same environmental conditions (phylogenetic effect).
- Differences that develop during the life of the individual organism (ontogenetic effect).
- Differences between skeletal units within a single organism (microarchitectural effect).
Relation between Temperature and K Mg/ Ca, K Sr/ Ca in inorganically precipitated calcite and aragonite:
- Sr concentration decreases with increasing temperature in both calcite and aragonite (Fig. 3-8).
- Mg concentration increases with increasing temperature.
Relation between Temperature and K Mg/ Ca, K Sr/ Ca in biologically precipitated calcite and aragonite:
- For biologically precipitated calcite and aragonite the trace element temperature relationship must be determined for each taxonomic group.
- Mg concentration correlates positively with temperature in many groups of organisms especially those having high Mg content (Fig. 3-9).
- Sr concentration sometimes correlates positively with temperature as in brachiopods and in some cases negatively as in echinoids.
It also has positive correlation in the calcite layer of Mytilus and negative correlation in the aragonite layer (Fig. 3-10).
Scleractinian corals show a negative correlation between Sr. concentration and temperature, which is very close to that found in inorganic precipitates of aragonite.
So, distribution coefficient for Mg and Sr can’t be used as paleotemperature indicator but can be only used to monitor the chemistry of the water in which the skeleton formed.
Trace chemistry and paleosalinity:
Trace chemistry can’t be used to determine the salinity of ancient environments because most Ca, Mg and Sr comes from the sea and in case of brackish water environment we will get incorrect results especially Mg/Ca and Sr/Ca ratios are nearly constant above a salinity of about 10%.
Other trace elements (Mn, Fe, Cd, B, Pb...):
Many other trace elements are found in modern and fossil skeletons such as Mn, Pb, Cd, B.... etc.
e.g.: Mn/ Ca and Fe/ Ca ratios are higher in fresh water than in marine water.
Cd/ Ca ratio is related to phosphate concentration and thus with productivity. It can be used to determine circulation pattern especially upwelling (and thus nutrient content) in the oceans.
Diagenetic effect:
As mentioned before, the older the fossil, the more likely it is to be altered chemically. On the other hand, the nature of the matrix in which the fossil is preserved may be of equal or greater importance. Preservation is most likely in a matrix that excludes contact between fossil and diagenetic fluid. There are few examples of Paleozoic fossil preserved in asphalt or fine, calcareous shaley matrix. Mg is less commonly preserved than Sr, this is because it is found in high Mg calcite, which is highly soluble.
Geologic application of trace chemistry:
1- Interpreting the trace chemistry of carbonate sediments, which compose mainly of skeletal fossils.
2- Interpreting diagenetic processes affecting carbonate sediments by comparing the original composition of fossil skeletons to that existed now.
3- Understanding geochemical cycle of elements.
4- Understanding chemical composition of ancient oceans.
5- Interpreting ancient environments such as distinguishing between freshwater fossils and marine fossils, and paleotemperature.
----------------------------------------------------------------------------------------------------------------
Oxygen isotopes as well as carbon isotopes of fossils can by used for paleoenvironmental interpretation, especially paleotemperature determination and paleosalinity (distinguishing marine water from fresh water).
Oxygen has three isotopes 16O, 17O and 18O. The first is more abundant in atmospheric oxygen followed by 18O and then 17O, which is least common.
Carbon has two isotopes; 12C which is more abundant than 13C. 14C occurs but in very small amounts.
Theory of applying isotopes:
Isotopes are forms of an element that differs only in the number of neutrons. They behave similarly but not identically in chemical reactions but differently in physical reactions. Both physical and chemical differences result in differences in the proportion of oxygen isotopes in skeletal material. The grater the difference in atomic weight (e.g. between 18O, 16O), the greater is the fractionation by physical and chemical processes.
Both oxygen and carbon atoms are present in carbonate skeletons in the CO32- ion. During skeletal formation these ions are in chemical equilibrium with oxygen and carbon present in the water as follow:
CO2 +H2O H2CO3 H + H CO3- 2H+ + CO32- ................(1)
A fractionation of isotopes occurs between water and carbonates in order to minimize the free energy of the system.
H218O + 1/3C16O32- H 216O + 1/3C18O32- .................(2)
Equilibrium K=[H 216O][ 3C18O32-]1/3/ [H218O][ 3C16O32- ]1/3
constant
Fractionation \a= (16O/18O) H2O X (18O /16O) CO32-
factor
a= (18O /16O) CO32-/ (18O /16O) H2O …..................(3)
OR: (18O /16O) CO32- = a (18O /16O) H2O ..................(4)
At 25°c, a =1.021 \ (18O) CO32- > (18O) H2O
· The same relationships also apply to carbon isotopes, which will fractionate between CO2 and CO32-.
In equation (4), as (18O /16O) H2O varies \(18O /16O) CO32- will also vary. This variation is usually caused by physical processes such as evaporation.
\ (H216O)L + (H218O)V (H218O)L + (H216O)V
a= (18O /16O)L/ (18O /16O)V
At 25°c a =1.008 \ (18O /16O)L >(18O /16O)V =0.8%
\18O in water is higher than 18O in vapor. This is because 16O is lighter and hence has higher vapor pressure and so evaporates faster.
Measurement of isotopic ratios:
Both oxygen and carbon isotopic ratios are measured by an instrument called mass spectrometer.
The sample used for measurements must be in the form of a gas like CO2 which is prepared from the sample by reaction with orthophosphoric acid.
6H+ + 2 PO43- + 3CaCO3 3CO2 + 3H2O + Ca2+ + 2 PO43-
Method
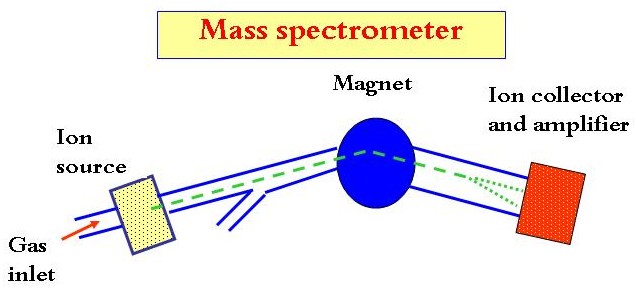
1- CO2 is introduced into the mass spectrometer through a gas inlet.
2- It enters the ion source when it is ionized into CO2+.
3- CO2 ion are accelerated and emerge as an ion beam that passes through a magnetic field.
4- The ions are deflected by an amount that is dependent on their mass, the lighter ions being deflected more than the heavier.
5- The ions discharge on the collectors and the amount of charge which is proportional to the ionic mass is measured.
· The ionic masses of special interest in oxygen and carbon isotopic studies are masses 44, 45,46.
Mass 44 consists of 12C 16O 16O.
Mass 45 consists of 13C 16O 16O.
Mass 46 consists of 12C 16O 18O.
18O /16O is obtained from mass 46/ mass 44.
13C/12C is obtained from mass 45/ mass 44.
Because there are other isotopes like 17O and also amount of the previously mentioned isotopes in the other masses, the measurement should be corrected with a standard.
\ d 18O (%) = (18O /16O) sample – (18O /16O) standard) X 1000
(18O /16O) standard
Del or delta
N. B. The standard used for paleotemperature determination is called PDB (Peedee Formation Belemnites), because it is prepared from specimens of Belemnintella americana. This species is composed of low Mg-calcite and so the specimens are well preserved (no diagenetic alteration).
Uses of isotopic techniques:
1- Paleotemperature determination.
2- Paleosalinity determination.
3- Correlation.
=============================================================
CHAPTER 3
Skeletal structure means the physical arrangement of mineral crystals within the skeleton. The study of this arrangement is useful in understanding the organism that formed the skeleton and also the influence of the environment.
Skeletal structure can be examined at two levels:
1- Finer level (ultrastructure): is the study of the individual crystallites making up the skeleton.
2- Coarser level (microarchitecture): is the study of the interrelationship of larger units within the skeleton.
Factors influencing skeletal structure:
1) Genetically controlled physiologic and biochemical factors.
2) Environmental factors.
Growth mechanism:
Skeletal structure is a function of the mechanism of growth of the skeleton. There are four basic methods of skeletal growth called accretion, molting, addition and modification.
1) Accretion :
Accretion is the building of skeleton by adding new material to the old part of skeleton. This happens in most organisms such as gastropods, corals, pelecypods …etc.
Organisms having this mode of growth all live in a tube or modified form of a tube. The tube may be coiled (gastropods, cephalopods, …etc), partially filled with structures such as septa and tabulae, allowing the organism to live only near the distal end of the tube (bryozoans and corals), partitioned into chambers with the animal occupying only the last chamber (cephalopods), doubled into two rapidly expanding tubes placed open end to open end and hinged to shut out the outside world (bivalves and brachiopods), grown together in colonies (corals and bryozoans) or equipped with a trapdoor (gastropods and cephalopods). The tube can grow in length and expand at its distal margins and it can increase the thickness of its walls by addition to the tube interior.
All shells growing by accretion have two-layered structure, one layer forms at the growing margin of the tube and the other forms at the inner surface of the tube.
2) Molting:
Molting is the periodical discarding of skeleton and formation of new larger one. This kind of growth is restricted to arthropods and probably is a major reason for their evolutionary success.
This method allows much more Variety of shapes because the skeleton can be essentially molded to the body. Its disadvantage is that the animal must periodically rebuild an entire new skeleton.
3) Addition:
This is the addition of skeletal elements to the skeleton such as addition of spicules to the spicular skeleton of sponge or addition of new plates to the echinoid test. The new plates are added at the apical region pushing down the old plates to the sides of the test. Because echinoid plates are surrounded by and permeated with living tissues, each plate grows also by accretion throughout the life of the specimen. Thus echinoderms grow by a combination of addition and accretion.
4) Modification:
This is the growth on all sides and even within the skeleton of vertebrate animals. Both the shapes and the size of the skeleton can change as needed.
___________________________________________________________
What is the relation between shell structure and environment?
Little is known about such relation in most invertebrates, something can be told about this relation in foraminifera, corals and mollusca.
There are three fundamental wall structures in foraminifera:
1- Agglutinated (arenaceous)
2- Hyaline
3- Porcellaneous.

The agglutinated structure consists of sediment grains acquired from the surrounding environment and embedded in a predominantly organic matrix.
The porcellaneous structure consist of randomly oriented elongate calcite crystals with a thin surface veneer of tangentially oriented crystals. It is nonporous and dense as compared to the hyaline structure.
The hyaline structure consists of somewhat larger elongate calcite crystals that are oriented perpendicular to the test surface. The hyaline wall is penetrated by numerous pores that parallel the crystals.
Relation to environment:
Foraminifera with agglutinated structure are especially common in low-salinity and relatively low temperature bays and estuaries. They are also relatively more abundant in deep water than shallow and in high latitudes than low.
Foraminifera with porcellaneous walls are especially abundant in shallow warm environment with normal to elevated salinity.
Foraminifera with hyaline structure are most common in areas with intermediate temperature and salinity conditions.
Explanation of variation in wall structure:
The reason for the presence of different wall structure in foraminifera is attributed to differing solubility of CaCO3. The solubility of CaCO3 varies with both temperature and salinity, cold and low-salinity waters are generally unsaturated with respect to CaCO3, and thus presumably, CaCO3 tests would be more difficult for the foraminifera to secrete, even with physiologic intervention.
High temperature and salinity waters are usually supersaturated with respect to CaCO3. The relatively massive, randomly nucleated porcellaneous structure can perhaps more readily secreted in such supersaturated waters.
In sea water saturated or only slightly supersaturated with respect to CaCO3, the porous but more highly organized hyaline structure is secreted.
The skeleton of modern scleractinian corals is composed of spherulitic clusters of aragonite needless. These clusters are arranged in vertical series forming elongate, fan-shaped trabeculae. The rugosa have a skeletal structure very similar to the scleractinia.
Relation to environment :
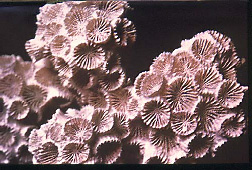 Environmental effects on
skeletal structure can be studied in terms of coral banding in the epitheca.
Environmental effects on
skeletal structure can be studied in terms of coral banding in the epitheca.
Banding occurs on two scales:
1) Constrictions and expansions with a spacing of about 1 cm (annual banding). The expansions presumably form during summer growth when conditions are especially favorable. Constrictions probably form during winter growth or during times of less favorable condition.
2) Fine ridge with a spacing of a few micrometers (daily banding). This is related to light intensity. Calcification in more rapid during the daylight hours than at night, presumably because of the effect of the symbiotic algae within the coral tissue. During the daylight hours these algae take up CO2 in their photosynthesis and thus raise the pH, aiding precipitation of CaCO3.
Significance of coral banding:
1- Determining the length of day in the geologic past by counting the number of daily growth lines between the annual expansion or constrictions.
Because of tidal friction the speed of rotation of the earth should be gradually diminishing and so the number of days pre years decreases.
In the Devonian, the number of days was calculated from coral bands and was 400 days.
2- Determining the age of the fossil coral.
3- Locating the latitudinal position of the area during that time.
This means that corals living near the equator should have uniform growth conditions and hence no banding. On the other hand, those living far from the equator where seasonality is pronounced, should have well developed banding. By this way we can determine if the area was near or far from the equator and so we can interpret continental drift during the past geologic time .
Banding in modern corals:
Banding in modern corals is difficult to recognize, so we use X-radiograph for studying such banding. Thin slices of coral colonies are prepared and then X-rays are transmitted through them onto X-ray sensitive film.
On the photograph, alternating dark and light bands appear. These appear to be related to variation in the density or compactness of the skeleton. The dense bands result from slower growth and are produced in late summer or early autumn. Some disturbance bands are also observed within the annual banding, these usually result from extremely low winter temperatures.
Factors controlling banding (growth rate):
1- Solar radiation
2- Suspended sediment
3- Temperature
4- Nutrient supply salinity
5- Wind and wave activity
Uses of banding in modern corals:
1- Determining the events occurring during the life of the coral .
For example, banding and 14C concentration were studied in modern corals. 14C was higher than expected in the early 18th century (a time characterized by slightly lower than normal temperature). This may be due to a slight decrease in solar activity at that time.
In the first half of the 20th century, 14C concentration decreased in corals, this is due to introduction of CO2 (containing no 14C) into the atmosphere from fossil fuel, resulting in the dilution of 14C in total atmosphere CO2.
In the second half of the 20th century, 14C concentration increased abruptly. This is due to production of 14C in atomic bomb testing.
90Sr concentration also increased in corals of the south pacific, this is also due to nuclear testing in that area. Its increase in corals of the Indian Ocean is due to transfer of water between the Pacific and Indian Ocean.
=============================================================
CHAPTER 4
Adaptive Functional Morphology
The physical, chemical and biological aspects of any environment change through time, as a result; organisms try to change themselves in order to better adapt with these changes. Such adaptive changes in fossil morphology can be used to detect ancient environments and this is what is called adaptive functional morphology.
There are three methods to study functional morphology:
1-Homology: studying functional morphology of a fossil by comparing it with its extant species (if present) or with a closely related living species.
Is it easy to detect all functions of morphological features?
In some cases it is easy, for example:
· The wedge shape of the clam for ease of burrowing.
· The claws of crabs for crushing prey.
· The stem of crinoids to elevate the calyx above sea floor.
In other cases, even in the living organisms, the function is not obvious, for example:
· Why are coral branches inclined into the current?
· Why does Nautilus have chambers?
· Why do the arms of crinoids form parabola-shaped nets?
2-Analogy: studying functional morphology of a fossil by comparing it with similar morphologies in extant organisms that are not closely related to the fossil.
For example: the spines in some extinct brachiopods functioned to prevent sinking into soft substrate just as do the spines in some modern bivalves.
3-Engineering or paradigm: studying functional morphology of a fossil through indirect engineering analysis of the function of a particular structure.
The use of fossil morphology to interpret ancient environments came from the observation of the fitness of living organisms to their environment.
Because organisms are adapted to their environment where the strong lives but the weak dies (natural selection) and this adaptation is in the form of morphological change, in this case and in the analysis of a fossil we can ask ourselves “For what environment is this morphology adapted? By answering this question, we can predict the environmental conditions under which the fossil lived”.
Reason for natural selection:
In any population, the full genetic content of the population will potentially be transmitted in unchanged proportion to the next generation unless outside forces operate. However, all the individuals in the population are not equally likely to survive because of genetic and corresponding physiologic and morphologic differences, which will cause them to be differentially adapted to the environment. Through the selection process only the best adapted individual form the breeding population that gives rise to the next generation and provides the genetic material that will determine the genetic and therefore, morphologic characteristics of that generation.
Selection is the “outside force” operating to change genetic composition. Selection results in changes in the phenotype (physiology- morphology- behavior) as well as genotype (genetic composition) of the organism.
Therefore, under stable environmental conditions, the genetic and morphologic composition will remain constant. But under changing environmental conditions, the genetic composition will change and hence adaptive functional morphology will exist to fit the new conditions.
Examples of functional morphology
Foraminifera
* Coiling direction in some species may be to the right (dextral) or to the left (sinistral). In Neogloboquadrina pachyderma coiling is sinistral when T< 9°c and is dextral when T> 15°c and mixed at T = 10-15°c.
The reason for that may be genetically linked with some physiologic features of foraminifera that has adaptive significance to temperature. The second reason is perhaps they are different species that have different temperature sensitivities. The third reason is perhaps one of the coiling direction is found in sexually produced form (microspheric (B) form) and the other in asexually produced form (megalospheric (A) form).
The porosity of the wall of planktonic forams varies with temperature. The more porous specimens, which are thus less dense, are formed under tropical condition where seawater is less dense and viscous than in colder climates.
Sponges
Sponges develop morphologic characters in response to environments especially turbulence.
Sponges living in quiet water have the problem of separating used water from new water. The sponge deals with this problem in two ways, the first: it has evolved a single osculum for expelling the used water at the upper end of the body. This allows maximum separation of the excurrent water from the incurrent water which enters at many separate ostia (pores) over the body.
The second way of adaptation is by increasing the distance to which the expelled water is removed from the body. This is done by increasing the pressure developed by the flagella and decreasing the size of the osculum (like a garden hose) in order to increase the velocity of excurrent water.
The vase-shape with narrow osculum is most suitable for that behavior in quiet water. Sponges living in turbulent water don’t have a vase-shape or narrow osculum.
Corals
1- In relation to depth:
The morphology of individual coral colonies varies with depth, some colonies have hemispherical forms (< 5 m), others columnar forms (5- 25m) and plate-like (>25m). This is due to differential growth rates controlled by light intensity.
2- In relation to substrate:
According to the type of substrate, the base of coral skeletons differ in its shape, which is, observed as sharp bending. Generally; corals growing in soft substrate tend to have a more conical base and forms living on firm substrate have a flatter base.
3- In relation to turbulence:
Massive and heavy branching corals are found in turbulent water whereas delicate branches are found in quiet water.
Massive forms from turbulent environments develop branching when transplanted in a quiet-water environment.
Heavy branching forms often have their branches oriented into the oncoming currents or waves.
Explanation for functional morphologic changes:
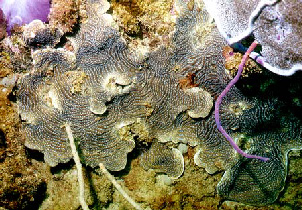
- In settings with no current, a vertical branching pattern minimizes mechanical stress on the skeleton due to gravity. In environments with strong currents, inclined orientation minimizes stress.
- Coral morphology may also be controlled by feeding characteristic. Sea fans or gorgonians (octacorals) are oriented with their planar surface perpendicular to current direction in order to get as much food as they can.
Bryozoans
Bryozoan animals show also a sort of functional morphology especially the fenestrate Paleozoic bryozoans. These colonies have the shape of the fenestrules as shown in the figure.
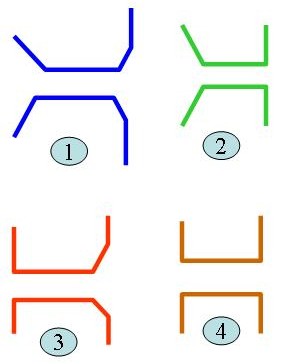
- In model (1), which is similar to actual bryozoan specimens, water-containing KMnO4 is allowed to flow at 2cm/sec.
- About 1/3 of KMnO4 passed through the model, the dyed water after passing the model eddied back forming a “curtain” The curtain persisted for 5-8 sec. Before dispersing.
- In model (3,4) a small portion of water passed and in models 2,4 no curtain was formed. Thus model 1 which is similar to actual fenestrate bryozoans is the most effective for feeding mechanism in a low-velocity, unidirectional currents.
Bivalves
In an extinct group of unusual morphology belonging to bivalves which is called rudistids, functional morphology can be studied in order to analyze such unusual morphology especially there is no close living representative.
The rudistids are inequivalved, sessile, epifaunal heterodont bivalves that flourished in the Tethyan seas during Cretaceous times. They became extinct at the end of the Cretaceous and left no close relatives either in phylogenetic or ecological terms.
The shell consists of a conical to cylindrical right valve and a small cap-like left valve.
They often grow cemented together in huge aggregations forming reefs or reef-like bodies.
Their sedimentary and stratigraphic setting indicates that they lived in shallow, clear water of normal salinity.
Functional morphology:

The body cavity in the conical valve is very small in comparison to the total volume of the shell. The earlier-formed portion of the shell is filled with tabulae which apparently served as successive platforms supporting the visceral mass that was suspended from the upper or left valve. The size of the body cavity was further reduced by: the thick shell, the massive tooth and socket structure and two or three infoldings or pillars of the valve wall.
The left valve formed a lid capping the opening of the cylinder of the right valve. It was equipped with massive teeth and myophores for muscle attachment that extended into the right valve. The left valve contains a complex system of ramifying canals that lead from the upper surface of the valve to the valve margin.
The growth lines in the shell structure indicate that the canals in the left valve must have been lined with living mantle tissues. These tissues were ciliated tubes that drew in water and suspended food from the outside and pumped it through the canals to the shell margin. The food particles were trapped on mucus strands and pass laterally along a trough-like structure on the right valve margin to the mouth.
The canal system served as a sieve device to exclude predators and sediment grains.
The current direction may have occasionally been reversed to clean out any sediment that would tend to clog the system.
The very short but strong muscles wouldn’t have allowed the valves to open more than a fraction of a millimeter. This is adequate for egress of the filtered water and waste products as well as more conventional filter feeding.
The rudistids were well adapted to life in high turbulence, shallow water environment. Their massive shells gave them stability in the turbulent environment.
They grew upward rapidly in competition with their neighbors for the food-laden water above; hence they developed their tall, cylindrical shape. The upward growth was facilitated by the development of tabulae in the right valve.
Finally, their feeding system was well adapted to a habitat with an abundance of suspended food.
Q. Why had the rudists died out although they well adapted to the environment?
Gastropods
Adaptive functional morphology can be studied by analogy between an extinct Paleozoic gastropod group and a distantly related modern gastropod group.
EX: G. Xenophora (the carrier shell).
This modern genus covers its shell with shells and shell fragments of other bivalves and gastropods. The function of this is for camouflage as the shell is difficult to see against a substrate containing many dead shells and shell fragments.
Xenophora also uses elongate shells (high-spired gastropods) to form spines or props around the margin of the shell slightly off the bottom. It also provides some protection and in addition, the animal can draw well back within its shell.
The animal hangs down from the shell and feeds on deposited organic material in the sediment.
The Xenophora mode of life requires a firm substrate in order to keep the shell suspended above the substrate, and an adequate supply of organic matter. They are mainly found in shallow, tropical, carbonate substrates, frequently in reef-associated environments.
Comparison:
There are some Paleozoic gastropods, which are similar to Xenophora in overall shape and presence of frill instead of props. By analogy to Xenophora, this gastropod must have lived on a firm substrate, feeding on deposited food.
An example of functional morphologic studies is genus Dendraster (sand dollar). This genus is disc-shaped with the mouth in the center and anus as well as apical system migrating towards the posterior. The apical system may be slightly off center or strongly off center…Why?
The genus can feed both on deposited and suspended food. During deposit feeding, Dendraster moves horizontally on the sea floor. During suspension feeding the animal partially buries the anterior part of the test in the sediment with the oral surface vertical or slightly inclined into the current.
In case of strong currents, the animal must bury a relatively large part of the anterior portion of the test. The eccentricity of the apical system allows the center of the apical system including the madreporite to be above the substrate so that respiration can take place and the water vascular system not become clogged with sediment. The stronger the currents, the more deeply the test must penetrate into the sediment for stability and thus the more eccentric the apical system must be to prevent its becoming buried.
So the morphologic features can be used as indicators of ancient current velocity.
================================================================
